Abstract
Adrenarche, the post-natal onset of adrenal production of DHEA (dehydroepiandrosterone) and its sulfate (DHEA-S) marks the 5-8 transition and the onset of middle childhood. However, the role of DHEA and/or DHEA-S in the cognitive, emotional and physical changes of the so-called 5-8 transition are not well-understood. Recent findings from the U.S. NIH Normal Brain and Development Study clearly demonstrate a role for DHEA in the development of the dorsolateral prefrontal cortex, associated with executive function, and the right temporal parietal junction, associated with mentalization. In addition, results from the Australian Childhood to Adolescence Transition study implicate DHEA in increased anxiety during middle childhood through increased connectivity of the amygdala with the anterior cingulate cortex and insula, a circuit that underlies both bodily perception and emotion. At the cellular level, DHEA acts on the sigma-1 receptor to promote both glutamatergic transmission and the production of BDNF (brain derived neurotrophic factor), which in turn promotes activity-dependent synaptic plasticity. The cerebellum has a high portion of glutamatergic neurons, hence DHEA may play a specific role in its development, including the promotion of cerebellar-cortico and cerebellar-limbic connectivity. Furthermore, such circuits have recently been implicated in the development of sensorimotor, cognitive and default mode systems during the 5-8 transition. Taken together these different lines of research suggest that adrenarche plays a key role in brain development and the changes in cognition, behavior and emotion that define the 5-8 transition and middle childhood. Research specifically considering the impact of DHEA on the development of the cerebellum in humans is called for.
Keywords
Adrenarche, DHEA, Middle childhood, Cerebellum, Glutamate
Introduction
Adrenarche, the onset of DHEAS production by the adrenal zona reticularis, is an important marker of middle childhood [1]. Middle childhood, referred to as the age of reason and responsibility is defined by a set of cognitive and behavioral changes, including mentalizing [2], cultural understanding [3], attachment [4], distancing from parents [1], and the emergence of narratively structured dreams [5]. Overshadowed by puberty, adrenarche has received little attention as a fundamental aspect of human neurological development despite the co-incidence of increasing levels of DHEAS, brain development and the 5-8 (years) behavioral transition.
However, recent findings from the U.S. NIH Normal Brain and Development Study (NBDS) have clearly demonstrated the impact of DHEA on brain development [6], while the Australian Childhood to Adolescence Transition Study (CATS) has implicated DHEA in the development of anxiety and social behavior [7] along with brain mechanisms underlying emotions [8]. At the same time, other research has shown a role for cerebellar-cortico connectivity in the development of brain systems underlying motor development, cognition and the default model network [9]. Separate research at the cellular level has implicated DHEA in glutamate transmission [10] and BDNF production [11] suggesting a role for DHEAS in synaptic plasticity associated with middle childhood. Here, I integrate these different lines of research to provide a clear picture of the role of DHEA in brain development during middle childhood and suggest directions for future research.
DHEA and Human Brain Development
Results from the NIH Normal Brain Development Study (NBDS) link DHEA to differences in regional cortical thickness across middle childhood. Most notably, Nguyen et al. [6] found a positive relationship between salivary DHEA and cortical thickness in the left dorsolateral prefrontal cortex (lDLPFC) from 4-8 years of age, and the right temporal parietal junction (rTPJ) from 7-12 years of age in a sample of 261 children. These results are directly related to key behavioral changes during middle childhood. The lDLPFC is central to the development of behavioral inhibition at the onset of middle childhood [12] and the rTPJ central to development of mentalizing from 7 to 12 [2]. As such they establish DHEA as central to middle childhood brain development.
Follow up results from the NBDS show an impact of DHEA on limbic-cortical connectivity underlying changes in memory and cognition as well. Nguyen et al. [13] report that salivary DHEA was related to covariance of the amygdala with the left occipital pole, the right somatosensory cortex, and the right anterior cingulate cortex (rACC), across the span of 6 to 22 years. In addition, DHEA was related to visual attention through covariation of the amygdala and the prefrontal cortex more globally. In other results, DHEA was related to working memory [14] and cognitive ability in boys [15] through its effects on cortico-hippocampal covariance. These findings were independent of age and pubertal hormones, indicating that the impact of DHEA on neural development continues even with the added impact of pubertal hormones on the brain.
At the same time, CATS has clearly implicated DHEAS in emotional and behavioral development during middle childhood for both sexes. A survey of over 1000 boys and girls, aged 8 & 9 years, showed positive relationship of DHEA-S (as distinct from DHEA) and conduct problems, emotional symptoms, and peer relationship problem among boys [7]. In contrast, for girls DHEA-S was positively related to peer conflict alone. Given CATS large sample size, these findings provide robust evidence that adrenarche is directly related to emotional and behavioral changes associated with middle childhood. They also suggest that the social implications of such changes may differ for boys and girls, with boys facing more general social adjustment and girls centered on their interactions with other girls.
CATS provides additional evidence for the impact of DHEA on brain regions underlying emotion and mood. Barandse et al. [16] report an indirect negative association of DHEAS with anxiety mediated by amygdala connectivity to insula and fusiform face area in girls, but not boys. A follow up study reported a positive indirect association of DHEA at time 1 with anxiety at time 2 mediated by amygdala connectivity with the frontal gyrus [16]. Using the same sample of children, Ellis et al. [17] also found a relationship between DHEA and hippocampal volume for both boys and girls, but no relationship between DHEA and depressive symptoms. These results are consistent with the NBDS findings in implicating DHEA in the development of the connections between the amygdala and insula, as well as the development of the hippocampus. (See Figure 1 for a diagrammatic representation of brain mechanism underlying the relationship with anxiety for girls).
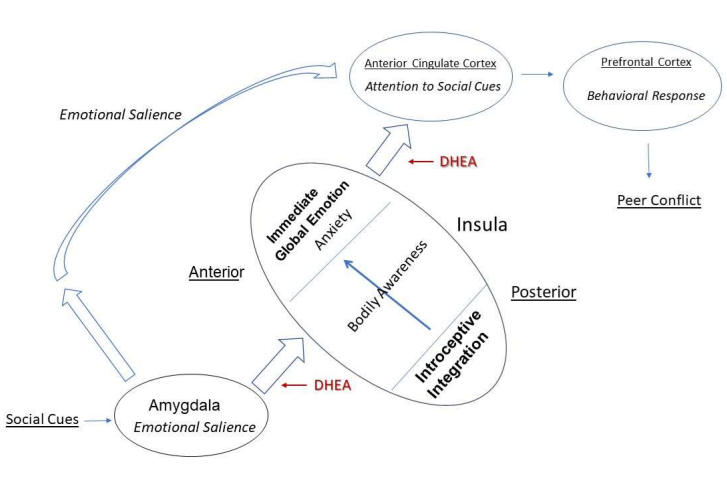
Figure 1: Schematic Representation of DHEA and Peer Conflict Among Girls. The anterior insula (AI) integrates bodily sensation from the posterior insula with emotional salience from the Amygdala to immediate global emotion, or bodily feeling in the moment. This information is conveyed to anterior cingulate cortex (ACC) where it is compared to direct information from the amygdala. If the signals are discrepant, a signal is sent to the prefrontal cortex to produce a behavioral response. With adrenarche, increasing levels of DHEA act to increase the connectivity of the insula with the amygdala and ACC. For girls this heightens the emotional salience of social cues from peers, leading to anxiety about their bodies.This anxiety promotes a behavioral response which leads to conflict with those peers.
Additional evidence for an impact of DHEA on the amygdala, insula and hippocampus comes from administration of exogenous DHEA to adults [18,19]. Sirpada et al. [18] reported that 400 mg of DHEA reduced the activity of the amygdala and hippocampus while increasing connectivity between the two regions. DHEA administration also enhanced activity in the rACC. Further results indicated that administration of DHEA reduced connectivity between the amygdala and insula [19]. At a functional level, DHEA administration reduced negative affect and memory. Together these results support a role for DHEA in the corticolimbic circuitry between the insula, anterior cingulate cortex, and anterior cingulate cortex underlying the emotional experience of the body [20], though the overall impact appears to differ between children and adults.
Other results from CATS draw attention to the potential impact of DHEA on myelinization. Barendse et al. [21] report a positive association of salivary DHEA with diffusivity in the corpus collosum, superior and posterior corona radiata and super longitudinal fasicularis among children, ages 9-10 years. However, a longitudinal follow up study [22] reported no relationship between changes in DHEA and changes in fiber density (FD) or fiber cross-section (FC) among children measured 1.5 years apart. An impact of DHEA on myelinization might be more apparent during puberty and young adulthood with the formation of neuronal networks that depend on well-insulated axonal transmission.
Cerebellar-Cortico Connectivity
In a study of the cerebellum across infants, children and adults, Kippling et al. [23] identified 7 cerebellar functional networks in infants, and 12 in children and adults. In infants, those 7 cerebellar networks were connected exclusively to the cortical sensorimotor network. In middle childhood, the 12 cerebellar networks were most strongly connected to sensorimotor, cognitive and default mode networks in the cortex, while for adults, coherence of cortico-cortical networks was most prominent.
Additional findings documented a relationship between lateral cerebellar white volumes and performance on the Stocking of Cambridge test, a standard cognitive measure [23].
In other words, the cerebellum not only plays a role in motor development, but during middle childhood its role expands to the development of cognition, and internal mentation as well. Given the cerebellum’s role in sequencing of internal mentation [24], and the role of the default mode in dreaming [25], connectivity with the default mode network suggests an important cerebellar role in the development of narrative dream structure mentioned at the outset of this article.
Molecular Mechanisms
Taken together, the findings discussed above suggest a clear role for DHEAS in brain development, including cortical thickness and connectivity underlying in cognition, emotion and behavior during middle childhood. However, DHEA’s impact on the molecular mechanisms underlying brain development have not been well elucidated. DHEA is known to have a variety of effects at the neuronal level [26], including promoting synaptogenesis [27], and neurotransmitter release [28], as well protecting against lipid peroxidation [29], all of which may promote neural connectivity. Here the author focus on DHEA’s actions on glutamate transmission and myelinization as one of the most promising potential mechanisms linking DHEA and neuroplasticity.
Neuronal processes
DHEA is known to act on a variety of non-genomic receptors, including sigma-1, GABA-A and glutamate [10]. Here the author suggests that DHEA impact on brain development involves two separate pathways; 1) altering glutamate transmission; 2) promoting BNDF release. In particular, the author considers the potential impact of DHEA on the sigma-1 receptor (SR-1). SR-1 is a complex molecule located on the mitochondrial membrane between the mitochondria and endoplasmic reticulum, with actions on multiple cellular processes, not all of which are fully understood [30]. Most importantly here, it is thought to translocate to the plasma membrane where it has an impact on NMDA receptors and BDNF release.
DHEA is known to has several effects on glutamate transmission, including enhancing the release of glutamate, as well as allosteric regulation of the glutamate NMDA receptor and inhibition of the GABA-A receptor [31]. DHEA has also been shown to promote activity of the astrocytic glutamate transporter (GLT-1) [32]. GLT-1 is involved in the removal of extra glutamate from the synaptic cleft reducing the potential for damage due to glutamate excitotoxicity (Figure 2).
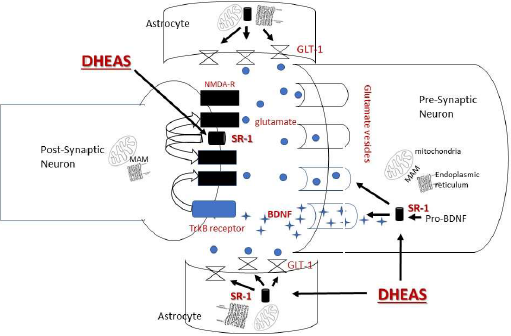
Figure 2: Proposed Impacts of DHEA on Glutamate Transmission. DHEA is suggested to alter glutamate transmission through its effects on the sigma-1 receptor (SR-1) in at least important three ways. In one pathway, DHEA acts on SR-1 which has translocated from the mitochondrial associated membrane (MAM) between the mitochondria and endoplasmic reticulum to the plasma membrane. At the plasma membrane, SR-1 alters the NMDR ionotropic receptor and modulates the impact of glutamate on the receptor itself. In another pathway, DHEA acts on SR-1 to change pro-BDNF to BDNF. BDNF promotes glutamate release pre-synaptically. Post-synaptically, BDNF acts at the TrKB receptor to alter the NMDA receptor. In addition, SR-1 acts on the glutamate transporter-1 (GLT-1) located in astrocytes to recycle glutamate thus reducing potential glutamate excitotoxicity
More recently, DHEA has been shown to promote BDNF [11]. BDNF itself promotes activity-dependent synaptogenesis through glutamate transmission, or more specifically by promoting the effects of SR-1 on the NMDA receptor [30]. SR-1 density in the human brain is highest in hippocampus, followed by the cerebellum and then cortical areas including the ACC [33]. As discussed above, connectivity of the amygdala to the hippocampus and the ACC has been linked to DHEA, which may reflect SR-1 potentiated release of BDNF as part of glutamate’s impact on activity-based synaptogenesis more generally.
Any impact of DHEA on glutamate and BDNF - related synaptic plasticity would be enhanced in the cerebellum. The cerebellum not only contains a high density of glutaminergic neurons, but expresses the GLUD2 gene which allows for faster glutamate cycling [34]. However, as far as the author is aware, there has been no work that considers the role of DHEA on cerebellar activity or its connection with cortical structures, let alone developmental changes in the cerebellum and cognition or emotion.
Myelinization
The co-incidence between on-going myelinization [35] and increasing DHEA level [36] both of which start around 6-7 years and continue into young adulthood suggests DHEA may promote myelinization of axons, an important aspect of longer-range neural connectivity. DHEA administration to rats has been shown to decrease brain lipid peroxidation as measured by malondialdehyde, thiobarbaturic acid reactive substance and hydroxy nonenal [37]. At the cellular level, DHEA has been shown to increase levels of the anti-oxidant manganese superoxide dismutase in mitochondria [38]. Thus, DHEA may help protect myelinization from the effects of reductive oxygen species associated with cellular energy production that is itself necessary for myelinization. However, there is currently little empirical data against which to consider this possibility.
Conclusion
The findings from the NBDS and CATS studies discussed here make clear adrenarche’s role in brain development and associated cognitive, emotional and behavioral changes during middle childhood. They implicate DHEA in the development of cortical thickness in regions central to executive function and mentalization during the 5-8 transition and middle childhood respectively. At the same time, they implicate DHEA in the development of anxiety as well as enhanced connectivity between the amygdala, insula and anterior cingulate circuit underlying emotion more generally. The author suggests that such developmental outcomes are related to DHEA’s effects on glutamate and BDNF which promote activity-dependent synaptic plasticity crucial for shaping brain development during middle childhood. In particular, the author point towards the development of the cerebellum and its connection to the cortex and limbic system during middle childhood.
Now that it is clear that DHEA is associated with cortical development, and the cerebellum with the development of cognition, emotion as well as motor control during the 5-8 transition, the next research step is to determine whether DHEA plays a role in the development of cerebellar-cortico and cerebellar-limbic connectivity middle childhood. Given an increasing focus on the impact of physical activity, obesity and stress on brain development during childhood, such research would help to delineate pathways by which these recognized factors alter adrenal function and impact brain development critical to the development of behavior, cognitive and emotion during middle childhood.
Acknowledgements
I thank Chawki Belhadi, Gillian Bentley and David Weissman and an anonymous reviewer for their comments on an earlier version of this manuscript.
References
2. Gweon H, Dodell‐Feder D, Bedny M, Saxe R. Theory of mind performance in children correlates with functional specialization of a brain region for thinking about thoughts. Child Development. 2012 Nov;83(6):1853-68.
3. Lancy DF, Grove MA. Getting noticed. Human Nature. 2011 Sep 1;22(3):281-302.
4. DelGiudice M. Middle childhood: An evolutionary-developmental synthesis. Child Development Perspectives. 2014;8:193-200.
5. Foulkes D. Children’s Dreaming and the Development of Consciousness. Harvard University Press, Cambridge, MA; 2002.
6. Nguyen TV, McCracken JT, Ducharme S, Cropp BF, Botteron KN, Evans AC, et al. Interactive effects of dehydroepiandrosterone and testosterone on cortical thickness during early brain development. Journal of Neuroscience. 2013 Jun 26;33(26):10840-8.
7. Mundy LK, Romaniuk H, Canterford L, Hearps S, Viner RM, Bayer JK, et al. Adrenarche and the emotional and behavioral problems of late childhood. Journal of Adolescent Health. 2015 Dec 1;57(6):608-16.
8. Barendse ME, Simmons JG, Byrne ML, Patton G, Mundy L, Olsson CA, et al. Associations between adrenarcheal hormones, amygdala functional connectivity and anxiety symptoms in children. Psychoneuroendocrinology. 2018 Nov 1;97:156-63.
9. Kipping J. Taun Ta, Fortier MW, Qia A. 2017. Asynchronous development of the cerebellum, cerebello-cortico and cortico-cortical functional networks in infancy, childhood and adulthood. Cerebral Cortex 27: 5170-5184.
10. Greaves RF, Wudy SA, Badoer E, Zacharin M, Hirst JJ, Quinn T, Walker DW. 2017. A tale of two steroids: The importance of the androgens DHEA and DHEAS for early neurodevelopment. J Steroid Biochem Mol Biol. 188:77-85.
11. Wu S, Ye M, Li Z, Bu S, Zhang Y. Long-term supplementation of dehydroepiandrosterone improved depressive-like behaviors by increasing BDNF expression in the hippocampus in ovariectomized rats. Heliyon. 2020 Oct 1;6(10):e05180.
12. Steinbeis N, Haushofer J, Fehr E, Singer T. Development of behavioral control and associated vmPFC–DLPFC connectivity explains children's increased resistance to temptation in intertemporal choice. Cerebral Cortex. 2016 Jan 1;26(1):32-42.
13. Nguyen TV, Gower P, Albaugh MD, Botteron KN, Hudziak JJ, Fonov VS, et al. The developmental relationship between DHEA and visual attention is mediated by structural plasticity of cortico-amygdalar networks. Psychoneuroendocrinology. 2016 Aug 1;70:122-33.
14. Nguyen TV, Wu M, Lew J, Albaugh MD, Botteron KN, Hudziak JJ, et al. Dehydroepiandrosterone impacts working memory by shaping cortico-hippocampal structural covariance during development. Psychoneuroendocrinology. 2017 Dec 1;86:110-21.
15. Farooqi NA, Scotti M, Yu A, Lew J, Monnier P, Botteron KN, et al. Sex‐specific contribution of DHEA‐cortisol ratio to prefrontal‐hippocampal structural development, cognitive abilities and personality traits. Journal of Neuroendocrinology. 2019 Feb;31(2):e12682.
16. Barendse ME, Simmons JG, Patton G, Mundy L, Byrne ML, Seal ML, et al. Adrenarcheal timing longitudinally predicts anxiety symptoms via amygdala connectivity during emotion processing. Journal of the American Academy of Child & Adolescent Psychiatry. 2020 Jun 1;59(6):739-48.
17. Ellis R, Fernandes A, Simmons JG, Mundy L, Patton G, Allen NB, et al. Relationships between adrenarcheal hormones, hippocampal volumes and depressive symptoms in children. Psychoneuroendocrinology. 2019 Jun 1;104:55-63.
18. Sripada RK, Marx CE, King AP, Rajaram N, Garfinkel SN, Abelson JL, et al. DHEA enhances emotion regulation neurocircuits and modulates memory for emotional stimuli. Neuropsychopharmacology. 2013 Aug;38(9):1798-807.
19. Sripada RK, Welsh RC, Marx CE, Liberzon I. The neurosteroids allopregnanolone and dehydroepiandrosterone modulate resting‐state amygdala connectivity. Human Brain Mapping. 2014 Jul;35(7):3249-61.
20. Campbell BC, Garcia JR. Neuroanthropology: evolution and emotional embodiment. Frontiers in Evolutionary Neuroscience. 2009 Nov 24;1:4.
21. Barendse ME, Simmons JG, Byrne ML, Seal ML, Patton G, Mundy L, et al. Brain structural connectivity during adrenarche: Associations between hormone levels and white matter microstructure. Psychoneuroendocrinology. 2018 Feb 1;88:70-7.
22. Barendse ME, Simmons JG, Smith RE, Seal ML, Whittle S. Adrenarcheal hormone-related development of white matter during late childhood. NeuroImage. 2020 Dec 1;223:117320.
23. Kipping JA, Xie Y, Qiu A. Cerebellar development and its mediation role in cognitive planning in childhood. Human Brain Mapping. 2018 Dec;39(12):5074-84.
24. Van Overwalle F, Ma Q, Heleven E. The posterior crus II cerebellum is specialized for social mentalizing and emotional self-experiences: a meta-analysis. Social Cognitive and Affective Neuroscience. 2020 Sep;15(9):905-28.
25. Fox KC, Nijeboer S, Solomonova E, Domhoff GW, Christoff K. Dreaming as mind wandering: evidence from functional neuroimaging and first-person content reports. Frontiers in Human Neuroscience. 2013 Jul 30;7:412.
26. Mellon SH. Neurosteroid regulation of central nervous system development. Pharmacology & Therapeutics. 2007 Oct 1;116(1):107-24.
27. MacLusky NJ, Hajszan T, Leranth C. Effects of dehydroepiandrosterone and flutamide on hippocampal CA1 spine synapse density in male and female rats: implications for the role of androgens in maintenance of hippocampal structure. Endocrinology. 2004 Sep 1;145(9):4154-61.
28. Dong LY, Cheng ZX, Fu YM, Wang ZM, Zhu YH, Sun JL, et al. Neurosteroid dehydroepiandrosterone sulfate enhances spontaneous glutamate release in rat prelimbic cortex through activation of dopamine D1 and sigma-1 receptor. Neuropharmacology. 2007 Mar 1;52(3):966-74.
29. Kumar P, Taha A, Sharma D, Kale RK, Baquer NZ. Effect of dehydroepiandrosterone (DHEA) on monoamine oxidase activity, lipid peroxidation and lipofuscin accumulation in aging rat brain regions. Biogerontology. 2008 Aug;9(4):235-46.
30. Ryskamp DA, Korban S, Zhemkov V, Kraskovskaya N, Bezprozvanny I. Neuronal sigma-1 receptors: signaling functions and protective roles in neurodegenerative diseases. Frontiers in Neuroscience. 2019 Aug 28;13:862.
31. Clark BJ, Prough RA, Klinge CM. Mechanisms of action of dehydroepiandrosterone. Vitamins and hormones. 2018 Jan 1;108:29-73.
32. Chen T, Tanaka M, Wang Y, Sha S, Furuya K, Chen L, et al. Neurosteroid dehydroepiandrosterone enhances activity and trafficking of astrocytic GLT-1 via σ1 receptor-mediated PKC activation in the hippocampal dentate gyrus of rats. Glia. 2017 Sep;65(9):1491-1503.
33. Sakata M, Kimura Y, Naganawa M, Oda K, Ishii K, Chihara K, et al. Mapping of human cerebral sigma1 receptors using positron emission tomography and [11C] SA4503. Neuroimage. 2007 Mar 1;35(1):1-8.
34. Burki F, Kaessmann H. Birth and adaptive evolution of a hominoid gene that supports high neurotransmitter flux. Nature Genetics. 2004 Oct;36(10):1061-3.
35. Miller DJ, Duka T, Stimpson CD, Schapiro SJ, Baze WB, McArthur MJ, et al. Prolonged myelination in human neocortical evolution. Proceedings of the National Academy of Sciences. 2012 Oct 9;109(41):16480-5.
36. Søeborg T, Frederiksen H, Mouritsen A, Johannsen TH, Main KM, Jørgensen N, et al. Sex, age, pubertal development and use of oral contraceptives in relation to serum concentrations of DHEA, DHEAS, 17α-hydroxyprogesterone, Δ4-androstenedione, testosterone and their ratios in children, adolescents and young adults. Clinica Chimica Acta. 2014 Nov 1;437:6-13.
37. Sinha N, Baquer NZ, Sharma D. Anti-lipidperoxidative role of exogenous dehydroepiendrosterone (DHEA) administration in normal ageing rat brain. Indian J Exp Biol. 43:420-4.
38. Grimm A, Schmitt K, Lang UE, Mensah-Nyagan AG, Eckert A. Improvement of neuronal bioenergetics by neurosteroids: implications for age-related neurodegenerative disorders. Biochimica et Biophysica Acta (BBA)-Molecular Basis of Disease. 2014 Dec 1;1842(12):2427-38