Abstract
In our recent published study entitled “Targeted Photodynamic Therapy using a Vectorized Photosensitizer coupled to a Folic Acid Analog induces Ovarian Tumor Cell Death and inhibits IL-6-mediated Inflammation”, we explore the effect of a new approach for the treatment of ovarian cancer. We found that photosensitizer coupled to an analog of folic acid (PSFAA) selectively accumulates in ovarian cancer cells and, upon light activation, induces an efficient photodynamic cytotoxicity. Moreover, we demonstrate that FRα-targeted PDT induces an inhibition of the pro-tumoral and immunosuppressive IL-6 cytokine secretion. The results support that FRα-targeted PDT not only eradicates residual ovarian tumor cells but could also modulate the immune microenvironment. In this commentary, we introduce more in-depth ovarian cancer immune background, summarize key findings of the published work, and discuss emerging questions and future research directions, focusing on FRα PDT-induced immune re-activation and its potential to re-sensitize ovarian cancer to immunotherapies.
FRα Targeted Photodynamic Therapy for Ovarian Cancer Cell Death
Epithelial ovarian cancer (EOC) is a highly aggressive malignant tumor. In 80% of cases, it is diagnosed at an advanced stage (Stage III/IV according to the International Federation of Gynecology and Obstetrics (FIGO)) when the disease has spread from the ovaries to the entire surface of the peritoneum and is therefore classified as peritoneal carcinomatosis of ovarian origin [1].This disease is characterized by extensive intraperitoneal dissemination and a high relapse rate, mainly due to the persistence of residual tumor lesions after initial treatment [1,2]. This is despite standard management combining cytoreductive surgery and platinum-based chemotherapy [1], as well as targeted therapies such as PARP inhibitors or antiangiogenic agents [2]. Ovarian cancer remains a highly lethal disease, with approximately 314,000 new cases and 207,000 deaths reported worldwide in 2020 [3]. The majority of patients relapse within two years, and chemoresistance is accompanied by an extremely poor prognosis [4,5]. Tumor progression occurs in a complex microenvironment that is both chronically inflammatory and deeply immunosuppressive, contributing to immune escape of the tumor and therapeutic failure [6].
In recent years, immune checkpoint inhibitors (ICIs) have significantly improved patient prognosis in a variety of solid tumors [7,8], but unfortunately, EOC has proven to be unresponsive to immunotherapies (particularly anti-PD1 and anti-PDL1). Indeed, pembrolizumab (anti-PD1) monotherapy showed an overall response rate of 8% in the KEYNOTE-100 study, the largest study to date [9,10]. This therapeutic failure can be explained by several reasons. First, EOC has a low tumor mutational burden (TMB), which limits the production of immunogenic neoantigens [11]. Indeed, the less mutations there are, the less likely the tumor is to produce neoepitopes, such as abnormal proteins, that can be recognized by the immune system. Secondly, ovarian cancer is characterized by a profoundly immunosuppressive tumor microenvironment, that limits the activation of the immune system [12]. Regulatory T cells (Tregs) play a central role in this process by participating in the establishment of immune tolerance. Notably, they secrete immunosuppressive cytokines such as IL-10 and TGF-β and exert a negative control on effector T cells and dendritic cells (DCs) via the expression of inhibitory molecules [13,14]. At the same time, CD8+ T cells infiltrating the tumor exhibit a phenotype of functional exhaustion due to tumor’s inhibitory checkpoints overexpression and an immunosuppressive cytokinic microenvironment, thereby compromising the effectiveness of the antitumor immune response [8,7]. Finally, tumor-associated macrophages (TAMs), which are predominantly polarized toward a pro-tumor and anti-inflammatory phenotype, exert powerful immunosuppressive functions, especially on dendritic cell function [15,16]. TAMs are also responsible for high secretion of pro-inflammatory and immunosuppressive cytokines, such as IL-6 and IL-10 [17,16]. For all these reasons, the EOC is classified as a “cold tumor” and therefore is not defined as immunogenic and is unresponsive to immunotherapy. This complexity highlights the need for detailed characterization and appropriate therapeutic targeting for the immune landscape of ovarian tumors.
Based on the observation that current therapeutic approaches struggle to effectively eliminate the disease, our study proposes a targeted photodynamic therapy (PDT) strategy based on a vectorized photosensitizer coupled to a folic acid analog, exploiting the overexpression of the folate receptor α (FRα) on the surface of ovarian tumor cells [18]. This approach led to the tumor selectivity of PDT, the limitation of off-target effects, and the optimization of ovarian tumor cells elimination [18]. PDT induces cell death mainly through the generation of reactive oxygen species (ROS) following the activation of a photosensitizer by appropriate light irradiation [19,20]. These ROS cause major oxidative damage to membranes, proteins, and DNA, triggering various cell death programs, including apoptosis and necrosis [19]. The induction of these deaths depends on the subcellular location of the photosensitizer, the light dose, and the intensity of the oxidative stress generated [19]. PDT-induced apoptosis causes the outer mitochondrial membrane permeabilization and leads to the release of pro-apoptotic factors, therefore promoting the recognition of dying cells by the immune system [21,22]. Similarly, PDT-induced lytic cell death, such as necrosis, is characterized by a rupture of the plasma membrane and release of intracellular contents, which may include enzymes and danger signals [22,23]. Necrotic cell death types are associated with a marked inflammatory response, which may contribute to the re-activate a strong immune response [24]. In this way, the identification of cell death type generated by our FRα-targeted PDT developed could be interesting, especially in the context of immunogenic cell death identification. Interestingly, we also show that FRα-targeted PDT induced significant and sustainable reduction of IL-6 secretion, a pivotal cytokine driving tumor progression and transformation while fostering an immunosuppressive tumor microenvironment [18], and dysregulated immune cell activation [25]. These results suggest that the impact of targeted PDT could go beyond simple local cytotoxicity and could trigger a more favorable reprogramming of the tumor microenvironment.
Hence, broader reflection on the immunomodulatory potential of FRα-targeted PDT and its ability to transform localized tumor destruction into a biologically relevant signal for antitumor immunity is not described and therefore needed, especially in the EOC, classically described as immunologically “cold”. In this way, in this commentary, we raise a central conceptual question:
“Can FRα-targeted PDT transform local tumor destruction into an immunogenic signal capable of attenuating the pro-tumor IL-6 axis and restoring effective anti-tumor immunity in ovarian cancer?”
PDT-Induced Immunogenic Cell Death
In recent advances, PDT has been described as causing “immunogenic” cell death (ICD) [21]. According to Galluzzi et al., ICD is defined as a form of regulated cell death sufficient to activate an adaptive immune response in an immunocompetent syngeneic host [26], i.e., cell death that is capable of inducing the maturation of dendritic cells, which can then effectively present tumor antigens and thereby lead to the initiation of a tumor-directed T-cell response. In this context, the oxidative stress induced by ROS generation during PDT causes damage to the endoplasmic reticulum, lysosomes, cell membranes, and mitochondria and promotes the exposure or release of danger associated molecular patterns (DAMPs), which play a central role in the activation of innate and adaptive immunity [26–28]. (Figure 1). In several cancers, different types of DAMPs are described to be associated with PDT induced immunogenic cell death.
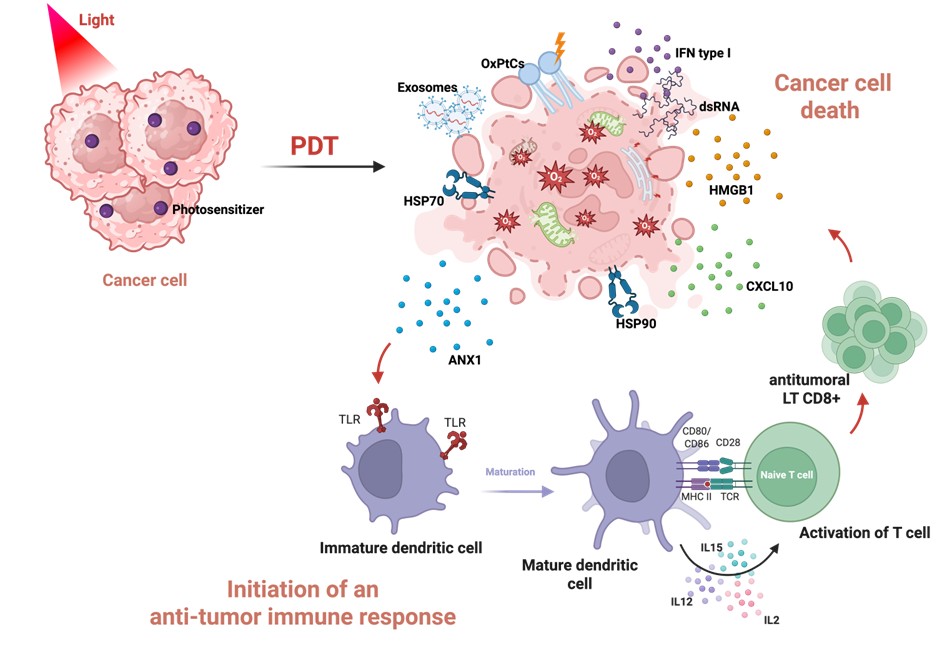
Figure 1. Overview of key molecular mechanisms of PDT-induced immunogenic cell death. Photodynamic therapy (PDT) induces immunogenic cell death associated with the release of danger signals (DAMPs), such as HSP70, calreticulin (CRT), high–mobility group box 1 (HMGB1), oxidized phosphatidylcholines (OxPtC), ANXA1. These signals are recognized by immature dendritic cells, notably via Toll-like receptors (TLRs), promoting their maturation. Mature dendritic cells subsequently present tumor antigens to naive T lymphocytes through MHC-TCR complexes and co-stimulatory molecules (CD80/CD86, CD28), while secreting immunostimulatory cytokines like IL2, IL15, and IL12, ultimately leading to an antitumor immune response. This PDT-induced immune reactivation could be investigated in the context of FRα-targeted PDT in epithelial ovarian cancer. (Figure created on Biorender and adapted from [54&55]).
First, there are membrane DAMPs, such as phosphatidylserine (PS), normally found in the inner leaflet of the plasma membrane, that is externalized during cell death induced by PDT, where it acts as an “eat-me” signal facilitating phagocytosis by immune cells [28]. Similarly, the exposure of heat shock proteins (HSP70, HSP90, GRP78/94) on the surface of dying cells has been reported to contribute to the absorption and presentation of tumor antigens [28–30]. Calreticulin (CRT), which can be exposed early on to the plasma membrane in response to endoplasmic reticulum stress, is considered one of the major signals of ICD and greatly enhances the immunogenicity of tumor cells treated with PDT [31,32]. This connection allows antigen-presenting cells (APCs) to migrate towards dying cells. Following oxidative stress induced by PDT, phosphatidylcholine (PtC) is oxidized to generate oxidized hosphatidylcholines (Ox-PtC) [28], which serve as recognition signals for phagocytic cells such as macrophages or dendritic cells. Among the membrane DAMPs, annexin A1, recognized by the FPR1 receptor, may also contribute to the guidance of antigen-presenting cells (APCs) to dying cells [28,29,33–35].
At the same time, several DAMPs can be released or secreted. High–mobility group box 1 (HMGB1), which is released upon loss of membrane integrity, has been shown to activate receptors such as Toll-like receptor 4 (TLR4) [31,32,36]. Other mediators, such as peroxiredoxin-1 (Prx1), may stimulate cytokine secretion via TLR4-dependent pathways, while ATP and UTP could play a central role in the recruitment and activation of antigen-presenting cells, as well as inflammasome activation [28,29].
In addition, cellular nucleic acids from dying cells, including cellular RNA, double-stranded RNA may be recognized by pattern recognition receptors such as TLR3 and TLR9, respectively, leading to the production of type I interferons. This production could promote the secretion of chemokines such as CXCL10, which is involved in the recruitment of effector T cells via CXCR3 [28,29,37].
Hence, the presence of such DAMPs types could be investigated after cell death induced by our FRα-targeted PDT in in vitro ovarian cancer cell models to validate its immunogenic potential.
Beyond these classic DAMPs, other signals remain less well characterized, including tumor cell-derived exosomes, which may exert immunomodulatory effects [25,28,38]. These extracellular vesicles could potentially be internalized by dendritic cells and contribute to antigen cross-presentation and T cell activation. In the context of ovarian cancer, a study by Baydoun et al. demonstrated that another FRα-targeted PDT induces the release of extracellular vesicles with immunoactivating properties, capable of increasing the proliferation of immune cells and thereby enhancing the antitumor response [25,28].
In line with these studies, a recent study conducted in our laboratory demonstrated that 5-ALA–based PDT in nasopharyngeal carcinoma profoundly alters the immunological properties of tumor-derived small extracellular vesicles (sEVs) [39]. While tumor-derived exosomes are typically associated with immunosuppressive functions, including impaired dendritic cell maturation and Treg recruitment [39,40]. PDT-modified sEVs displayed enhanced immunostimulatory properties. sEVs isolated from PDT-treated tumor cells induced a marked increase in PBMC proliferation and were enriched in double-stranded DNA [39]. These findings suggest that PDT reshapes the extracellular vesicle-mediated dialogue within the tumor microenvironment, potentially favoring immune activation.
Altogether, these signals provide a mechanistic framework linking innate and adaptive immunity, promoting dendritic cell maturation, cross-presentation of tumor antigens, and activation of cytotoxic T cells [29,41] (Figure 1). This process could thus counterbalance the suppressive state acquired by DCs during tumor progression and restore their ability to initiate an effective cytotoxic T cell response in EOC [32,42]. However, in the context of FRα-targeted PDT developed in our study, the induction of these key ICD hallmarks, including calreticulin exposure, HMGB1 and ATP release, and dendritic cell activation, has not yet been directly demonstrated. Therefore, while our current findings, including tumor cell death, reduced IL-6 secretion, and the absence of overt immunosuppressive effects on immune cell proliferation, suggest a potential immunomodulatory role, additional experimental validation is required to formally establish the capacity of this approach to induce ICD. The evaluation of such DAMPs with the FRα-targeted PDT developed in our study [18] thus represents a critical first step toward determining whether FRα-targeted PDT can effectively trigger immunogenic cell death.
The Role of Dendritic Cells in PDT-Induced Immune Activation
The release of DAMPs following PDT-induced tumor cell death, does not guarantee the establishment of a lasting antitumor immune response, as the effectiveness of these immunogenic signals depends on their processing by dendritic cells (DC), the central orchestrators of antitumor immunity [29,43]. The concept of the cancer-immunity cycle, described by Mellman et al., further reinforces the central role of dendritic cells in this antitumor immunity [44]. They showed that T cells activation in lymph nodes is highly dependent on DC throughout the antitumor response, but also that DC are now considered essential within the tumor itself to support the activity of infiltrating T cells [45]. Thus, DC maturation and activation appear to be central elements in ensuring an effective and sustained antitumor immune response (Figure 1).
In ovarian cancer, the tumor progression is accompanied by a profound reprogramming of dendritic cells (DCs), which causes the shift from effective antitumor immunity to a state of immune escape [42]. It has been shown that DCs from advanced stages of tumor growth have significantly lower levels of activation of the major histocompatibility complex class II (MHC II) and the costimulatory co-receptor CD40, impaired IL-12 production, and a diminished capacity to activate CD8+ T cells, ultimately promoting immune escape [42,46,47]. Thus, tumor progression in ovarian cancer is not solely based on mechanisms intrinsic to cancer cells, but also relies on a suppressive reprogramming of DCs, preventing their maturation, T cell activation, and expression of their effector function [48,49]. Therefore, beyond inducing immunogenic signals, an effective therapeutic strategy must also restore DC functionality within the tumor microenvironment.
In this context, several studies suggest that PDT has a major impact on DC maturation and migration, thereby contributing to the reactivation of antitumor immunity. In ovarian cancer, Baydoun et al. showed that PDT targeting the folate receptor induces a significant increase in PBMC proliferation in vitro and in vivo, indicating a generally immunostimulatory effect post-PDT [50,51]. Although DCs were not directly analyzed in these studies, these results may indirectly suggest DC activation. Similarly, studies of Gollnick et al. and Ji et al. demonstrated that exposure of DCs to DAMPs and antigens released by breast and skin cancer cells killed by PDT induces their functional maturation, characterized by the expression of MHC II markers and an increase in IL-12 production; [52]. Beyond maturation, PDT can also promote DC migration. Trempolec et al. highlights a significant increase in the expression of CCR7 on DC, favoring their migration [41,53].
These characteristics are important for the establishment of an effective adaptive immune response, even in tumors that are initially not very immunogenic. In the context of EOC, it seems crucial to examine not only the ability of PDT to induce immunogenic signals, but above all its ability to functionally reprogram dendritic cells that have become dysfunctional in the advanced stages of the disease. We hypothesize that FRα-targeted PDT could generate DAMPs that could rescue the dysfunctional phenotype of ovarian tumor-associated DCs by restoring MHC II and CD40 expression and promoting IL-12 secretion. This could be verified in vitro by co-culturing dendritic cells isolated from the ascites of patients with EOC with the culture medium from ovarian cancer cells treated with PDT or with in vivo models of humanized EOC [50]. Thus, careful evaluation of the impact of FRα-targeted PDT proposed in our study [18,25] on DC maturation, antigen-presenting capacity, and immunostimulatory function appears to be a future key step in considering this targeted PDT as an immune reprogramming strategy in ovarian cancer (Figure 1).
Modulation of the Cytokine Microenvironment by PDT to Re-Polarize the Immune Response
In addition to the potent stimulation of an immunogenic cell death, PDT could act indirectly on the cytokine microenvironment that led the immunosuppressive capacity of ovarian carcinomatosis.
In EOC metastatic states, ascite is a central biological compartment of the tumor microenvironment, characterized by a high concentration of pro-inflammatory and immunosuppressive cytokines. Ascites is particularly enriched in pro-inflammatory cytokines such as IL-6, IL-8, TNF-α, and IL-1β, which contribute to chronic inflammation, tumor survival, and peritoneal dissemination [56–58]. At the same time, it contains high levels of immunosuppressive cytokines and mediators, including IL-10, TGF-β [59], and VEGF, promoting the expansion of regulatory T cells, the polarization of M2-type tumor-associated macrophages [60], and the inhibition of dendritic cell maturation and function [6]. This inflammatory paradox, combining persistent inflammation and immune suppression, leads to a state of dysfunctional immunity in ascites, limiting the activation of effector T cells and contributing to the failure of antitumor responses and immunotherapy strategies [59,60].
PDT could act indirectly on this inflammatory microenvironment by regulating the IL-6 axis, whose chronic activation is strongly implicated in DC dysregulation and CD8+ and NK immunosuppression in EOCs [61–63]. The decrease in IL-6 secretion observed in the post-PDT secretome of the OVCAR3 and SKOV3 cell lines in vitro in the study by Boidin et al. [18] suggests a downregulation of this pro-tumor pathway, which could lift the inhibition of dendritic cell maturation and T-cell effector function (Figure 2). However, extensive data on the effect of such PDT in humanized in vivo EOC models are needed to confirm these in vitro observations. To support this hypothesis, a recent contribution has been made by the study by Amer et al. (2025), which demonstrates that IL-6 is a decisive factor in defining the “hot” or “cold” nature of the ovarian tumor microenvironment [61]. The authors show that high levels of IL-6 shape a profoundly immunosuppressive TME, maintaining chronic deleterious inflammation and promoting resistance to immune surveillance. Amer et al. also suggest that inhibiting IL-6, particularly in combination with checkpoint inhibitors (anti-PD-1/PD-L1), could help overcome this immunosuppressive barrier and promote the transition toward a more immunotherapy-responsive tumor phenotype.
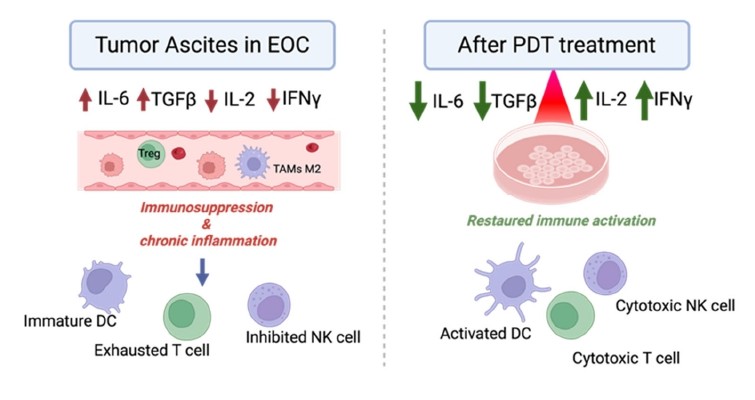
Figure 2. Modulation of the cytokine microenvironment by photodynamic therapy in ovarian cancer. FRα-targeted PDT modulates the ascites microenvironment in EOC in vitro, reducing immune suppressive cytokines IL-6 and TGF-β and increasing immunomodulatory cytokine IL-2 and IFN-γ. These changes may be associated with the lifting of immunosuppression and an activation of DCs, cytotoxic T lymphocytes, and NK cells in EOC [18,50,51]. (Figure created on Biorender).
Furthermore, Baydoun et al. show in vitro and in vivo that FRα-targeted PDT in ovarian cancer models can rebalance the cytokine inflammatory microenvironment. They demonstrate that PDT can reduce TGF-β, an immunosuppressive cytokine responsible for the expansion of regulatory T cells and the inhibition of effector T cells. Conversely, PDT promotes an increase in IL-2 and IFN-γ, two key cytokines in antitumor immunity, with IL-2 supporting the proliferation and survival of activated T lymphocytes and IFN-γ enhancing CD8+ cytotoxicity and Th1 polarization. Taken together, these results show that PDT could modulate the tumor microenvironment and could induce a reactivation of the immune system in EOC [25,50].
Future Therapeutic Direction
Thus, while the absence of an immunosuppressive effect on immune cell proliferation observed in our article is an encouraging sign toward immunomodulation, it does not in itself allow us to conclude that there is antitumor immune activation. To fully demonstrate the immunogenicity of FRα-targeted PDT, additional analyses would be necessary, including in vitro post-PDT DAMPs release analysis, dendritic cell maturation with PDT conditioned media, and co-culture with CD8 T cells to verify the modulation of immune system reactivation. Furthermore, in vivo validation using humanized murine models would be essential to determine whether these effects translate into a reprogramming of tumor microenvironment and a durable antitumor immune response.
Therapeutically, the combination of targeted PDT with immunotherapies, particularly ICIs, appears particularly promising [64]. By inducing immunogenic cell death and relieving IL-6–mediated immunosuppression, FRα-targeted PDT could create an environment permissive to CD8+ T cell infiltration and function, thereby rendering the tumor more sensitive to PD-1/PD-L1 blockade. In this context, FRα-targeted PDT could transform the “cold” microenvironment of EOC into “hot” one and improve its sensitivity to checkpoint blockade by restoring effective antitumor immune responses [65]. Hence, FRα-targeted PDT could contribute to the eradication of residual metastases after surgery of EOC, which could reduce the risk of relapses [66].
Conclusions
In conclusion, the FRα-targeted PDT is not limited to a localized tumor destruction strategy but emerge as a potential tool for immuno-inflammatory reprogramming of the peritoneal carcinomatosis microenvironment in ovarian cancer. By combining tumor selectivity and the induction of immunogenic cell death and cytokines modulations, FRα targeted PDT could open up innovative prospects for limiting peritoneal recurrence and improving the prognosis of patients with advanced EOC by favoring an immune “hot” microenvironment, potentially more sensitive to immunotherapies.
Acknowledgment
This research was supported by SATT Nord de France (PRODYNOV valorization contract). The University of Lille grants Nadia Hage Ali.
Conflicts of Interest
The authors declare no conflicts of interest.
References
2. Bhatt A, Sinukumar S, Kepenekian V, Kammar P, Mehta S, Shaikh S, et al. Platinum resistant recurrence and early recurrence in a multi-centre cohort of patients undergoing interval cytoreductive surgery for advanced epithelial ovarian cancer. Front Oncol. 2022 Sep 2;12:951419.
3. Cabasag CJ, Fagan PJ, Ferlay J, Vignat J, Laversanne M, Liu L, et al. Ovarian cancer today and tomorrow: A global assessment by world region and Human Development Index using GLOBOCAN 2020. Int J Cancer. 2022 Nov 1;151(9):1535–41.
4. Summey R, Uyar D. Ovarian cancer resistance to PARPi and platinum-containing chemotherapy. Cancer Drug Resist. 2022 Jun 22;5(3):637–46.
5. Freimund AE, Beach JA, Christie EL, Bowtell DDL. Mechanisms of Drug Resistance in High-Grade Serous Ovarian Cancer. Hematol Oncol Clin North Am. 2018 Dec;32(6):983–96.
6. Almeida-Nunes DL, Mendes-Frias A, Silvestre R, Dinis-Oliveira RJ, Ricardo S. Immune Tumor Microenvironment in Ovarian Cancer Ascites. Int J Mol Sci. 2022 Sep 14;23(18):10692.
7. Yan Y, Lu J, Luo H, Wang Z, Xu K, Wang L, et al. Decoding immune low-response states in ovarian cancer: insights from single-cell and spatial transcriptomics for precision immunotherapy. Front Immunol. 2025 Sep 19;16:1667464.
8. Wang W, Zou W, Liu JR. Tumor-infiltrating T cells in epithelial ovarian cancer: predictors of prognosis and biological basis of immunotherapy. Gynecol Oncol. 2018 Oct;151(1):1–3.
9. Ghisoni E, Morotti M, Sarivalasis A, Grimm AJ, Kandalaft L, Laniti DD, et al. Immunotherapy for ovarian cancer: towards a tailored immunophenotype-based approach. Nat Rev Clin Oncol. 2024 Nov;21(11):801–17.
10. Matulonis UA, Shapira-Frommer R, Santin AD, Lisyanskaya AS, Pignata S, Vergote I, et al. Antitumor activity and safety of pembrolizumab in patients with advanced recurrent ovarian cancer: results from the phase II KEYNOTE-100 study. Ann Oncol. 2019 Jul 1;30(7):1080–7.
11. Morand S, Devanaboyina M, Staats H, Stanbery L, Nemunaitis J. Ovarian Cancer Immunotherapy and Personalized Medicine. Int J Mol Sci. 2021 Jun 18;22(12):6532Morand S, Devanaboyina M, Staats H, Stanbery L, Nemunaitis J. Ovarian Cancer Immunotherapy and Personalized Medicine. Int J Mol Sci. 2021 Jun 18;22(12):6532.
12. Blanc-Durand F, Clemence Wei Xian L, Tan DSP. Targeting the immune microenvironment for ovarian cancer therapy. Front Immunol. 2023 Dec 18;14:1328651.
13. Moi D, Zeng B, Minnie SA, Bhatt R, Wood J, Sester DP, et al. Multiparametric flow cytometry to characterize vaccine-induced polyfunctional T cell responses and T cell/NK cell exhaustion and memory phenotypes in mouse immuno-oncology models. Front Immunol. 2023 Apr 6;14:1127896.
14. Yu L, Yang F, Zhang F, Guo D, Li L, Wang X, et al. CD69 enhances immunosuppressive function of regulatory T-cells and attenuates colitis by prompting IL-10 production. Cell Death Dis. 2018 Sep 5;9(9):905.
15. Nowak M, Klink M. The Role of Tumor-Associated Macrophages in the Progression and Chemoresistance of Ovarian Cancer. Cells. 2020 May 22;9(5):1299.
16. Wang Y, Ma C, Li X, Yang F, Wang N, Ji G, et al. Unraveling the role of M2 TAMs in ovarian cancer dynamics: a systematic review. J Transl Med. 2025 Jun 3;23(1):623.
17. Pan Y, Yu Y, Wang X, Zhang T. Tumor-Associated Macrophages in Tumor Immunity. Front Immunol. 2020 Dec 3;11:583084.
18. Boidin L, Moinard M, Moussaron A, Merlier M, Moralès O, Grolez GP, et al. Targeted Photodynamic Therapy using a Vectorized Photosensitizer coupled to Folic Acid Analog induces Ovarian Tumor Cell Death and inhibits IL-6-mediated Inflammation. J Control Release. 2024 Jul;371:351–70.
19. Kessel D. Death Pathways Associated with Photodynamic Therapy. Photochem Photobiol. 2021 Sep;97(5):1101–3.
20. van Straten D, Mashayekhi V, de Bruijn HS, Oliveira S, Robinson DJ. Oncologic Photodynamic Therapy: Basic Principles, Current Clinical Status and Future Directions. Cancers (Basel). 2017 Feb 18;9(2):19.
21. Chou W, Sun T, Peng N, Wang Z, Chen D, Qiu H, et al. Photodynamic Therapy-Induced Anti-Tumor Immunity: Influence Factors and Synergistic Enhancement Strategies. Pharmaceutics. 2023 Nov 11;15(11):2617.
22. Tan L, Shen X, He Z, Lu Y. The Role of Photodynamic Therapy in Triggering Cell Death and Facilitating Antitumor Immunology. Front Oncol. 2022 May 27;12:863107.
23. Gunaydin G, Gedik ME, Ayan S. Photodynamic Therapy-Current Limitations and Novel Approaches. Front Chem. 2021 Jun 10;9:691697.
24. Choi M, Shin J, Lee CE, Chung JY, Kim M, Yan X, Yang WH, Cha JH. Immunogenic cell death in cancer immunotherapy. BMB Rep. 2023 May;56(5):275–86.
25. Baydoun M, Moralès O, Frochot C, Ludovic C, Leroux B, Thecua E, et al. Photodynamic Therapy Using a New Folate Receptor-Targeted Photosensitizer on Peritoneal Ovarian Cancer Cells Induces the Release of Extracellular Vesicles with Immunoactivating Properties. J Clin Med. 2020 Apr 21;9(4):1185.
26. Galluzzi L, Vitale I, Warren S, Adjemian S, Agostinis P, Martinez AB, et al. Consensus guidelines for the definition, detection and interpretation of immunogenic cell death. J Immunother Cancer. 2020 Mar;8(1):e000337.
27. Li H, Shen J, Zheng C, Zhu P, Yang H, Huang Y, et al. Cell death: The underlying mechanisms of photodynamic therapy for skin diseases. Interdisciplinary Medicine. 2025 Mar;3(2):e20240057.
28. Garg AD, Krysko DV, Vandenabeele P, Agostinis P. DAMPs and PDT-mediated photo-oxidative stress: exploring the unknown. Photochem Photobiol Sci. 2011 May;10(5):670–80.
29. Galluzzi L, Buqué A, Kepp O, Zitvogel L, Kroemer G. Immunogenic cell death in cancer and infectious disease. Nat Rev Immunol. 2017 Feb;17(2):97–111.
30. Korbelik M, Sun J, Cecic I. Photodynamic therapy-induced cell surface expression and release of heat shock proteins: relevance for tumor response. Cancer Res. 2005 Feb 1;65(3):1018–26.
31. Gameiro SR, Jammeh ML, Wattenberg MM, Tsang KY, Ferrone S, Hodge JW. Radiation-induced immunogenic modulation of tumor enhances antigen processing and calreticulin exposure, resulting in enhanced T-cell killing. Oncotarget. 2014 Jan 30;5(2):403–16.
32. Garg AD, Nowis D, Golab J, Vandenabeele P, Krysko DV, Agostinis P. Immunogenic cell death, DAMPs and anticancer therapeutics: an emerging amalgamation. Biochim Biophys Acta. 2010 Jan;1805(1):53–71.
33. Vacchelli E, Ma Y, Baracco EE, Sistigu A, Enot DP, Pietrocola F, et al. Chemotherapy-induced antitumor immunity requires formyl peptide receptor 1. Science. 2015 Nov 20;350(6263):972–8.
34. Michaud M, Martins I, Sukkurwala AQ, Adjemian S, Ma Y, Pellegatti P, et al. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice. Science. 2011 Dec 16;334(6062):1573–7.
35. Garg AD, Dudek AM, Ferreira GB, Verfaillie T, Vandenabeele P, Krysko DV, et al. ROS-induced autophagy in cancer cells assists in evasion from determinants of immunogenic cell death. Autophagy. 2013 Sep;9(9):1292–307.
36. Sims GP, Rowe DC, Rietdijk ST, Herbst R, Coyle AJ. HMGB1 and RAGE in inflammation and cancer. Annu Rev Immunol. 2010;28:367–88.
37. Sistigu A, Yamazaki T, Vacchelli E, Chaba K, Enot DP, Adam J, et al. Cancer cell-autonomous contribution of type I interferon signaling to the efficacy of chemotherapy. Nat Med. 2014 Nov;20(11):1301–9.
38. Wu Q, Zhang H, Sun S, Wang L, Sun S. Extracellular vesicles and immunogenic stress in cancer. Cell Death Dis. 2021 Oct 1;12(10):894.
39. Trioën C, Soulier T, Massoud J, Bouchez C, Stoup N, Lefebvre A, et al. Efficient 5-ALA-photodynamic therapy in nasopharyngeal carcinoma induces an immunoactivation mediated by tumoral extracellular vesicles and associated with immunogenic cell death. Cancer Commun (Lond). 2025 Apr;45(4):401–5.
40. Olejarz W, Dominiak A, Żołnierzak A, Kubiak-Tomaszewska G, Lorenc T. Tumor-Derived Exosomes in Immunosuppression and Immunotherapy. J Immunol Res. 2020 May 22;2020:6272498.
41. Trempolec N, Doix B, Degavre C, Brusa D, Bouzin C, Riant O, et al. Photodynamic Therapy-Based Dendritic Cell Vaccination Suited to Treat Peritoneal Mesothelioma. Cancers (Basel). 2020 Feb 27;12(3):545.
42. Scarlett UK, Rutkowski MR, Rauwerdink AM, Fields J, Escovar-Fadul X, Baird J, et al. Ovarian cancer progression is controlled by phenotypic changes in dendritic cells. J Exp Med. 2012 Mar 12;209(3):495–506.
43. Kroemer G, Galluzzi L, Kepp O, Zitvogel L. Immunogenic cell death in cancer therapy. Annu Rev Immunol. 2013;31:51–72.
44. Mellman I, Chen DS, Powles T, Turley SJ. The cancer-immunity cycle: Indication, genotype, and immunotype. Immunity. 2023 Oct 10;56(10):2188–205.
45. Pittet MJ, Di Pilato M, Garris C, Mempel TR. Dendritic cells as shepherds of T cell immunity in cancer. Immunity. 2023 Oct 10;56(10):2218–30.
46. Huarte E, Cubillos-Ruiz JR, Nesbeth YC, Scarlett UK, Martinez DG, Buckanovich RJ, Benencia F, Stan RV, Keler T, Sarobe P, Sentman CL, Conejo-Garcia JR. Depletion of dendritic cells delays ovarian cancer progression by boosting antitumor immunity. Cancer Res. 2008 Sep 15;68(18):7684–91.
47. Krempski J, Karyampudi L, Behrens MD, Erskine CL, Hartmann L, Dong H, et al. Tumor-infiltrating programmed death receptor-1+ dendritic cells mediate immune suppression in ovarian cancer. J Immunol. 2011 Jun 15;186(12):6905–13.
48. Scarlett UK, Cubillos-Ruiz JR, Nesbeth YC, Martinez DG, Engle X, Gewirtz AT, et al. In situ stimulation of CD40 and Toll-like receptor 3 transforms ovarian cancer-infiltrating dendritic cells from immunosuppressive to immunostimulatory cells. Cancer Res. 2009 Sep 15;69(18):7329–37.
49. Caro AA, Deschoemaeker S, Allonsius L, Coosemans A, Laoui D. Dendritic Cell Vaccines: A Promising Approach in the Fight against Ovarian Cancer. Cancers (Basel). 2022 Aug 21;14(16):4037.
50. Baydoun M, Boidin L, Leroux B, Vignion-Dewalle AS, Quilbe A, Grolez GP, et al. Folate Receptor Targeted Photodynamic Therapy: A Novel Way to Stimulate Anti-Tumor Immune Response in Intraperitoneal Ovarian Cancer. Int J Mol Sci. 2023 Jul 10;24(14):11288.
51. Baydoun M, Moralès O, Frochot C, Ludovic C, Leroux B, Thecua E, et al. Photodynamic Therapy Using a New Folate Receptor-Targeted Photosensitizer on Peritoneal Ovarian Cancer Cells Induces the Release of Extracellular Vesicles with Immunoactivating Properties. J Clin Med. 2020 Apr 21;9(4):1185.
52. Ji J, Fan Z, Zhou F, Wang X, Shi L, Zhang H, Wang P, Yang D, Zhang L, Chen WR, Wang X. Improvement of DC vaccine with ALA-PDT induced immunogenic apoptotic cells for skin squamous cell carcinoma. Oncotarget. 2015 Jul 10;6(19):17135–46.
53. González FE, Ortiz C, Reyes M, Dutzan N, Patel V, Pereda C, et al. Melanoma cell lysate induces CCR7 expression and in vivo migration to draining lymph nodes of therapeutic human dendritic cells. Immunology. 2014 Jul;142(3):396–405.
54. Thiruppathi J, Vijayan V, Park IK, Lee SE, Rhee JH. Enhancing cancer immunotherapy with photodynamic therapy and nanoparticle: making tumor microenvironment hotter to make immunotherapeutic work better. Front Immunol. 2024 Apr 5;15:1375767.
55. Falk-Mahapatra R, Gollnick SO. Photodynamic Therapy and Immunity: An Update. Photochem Photobiol. 2020 May;96(3):550–59.
56. Lane D, Matte I, Rancourt C, Piché A. Prognostic significance of IL-6 and IL-8 ascites levels in ovarian cancer patients. BMC Cancer. 2011 May 30;11:210.
57. Kolomeyevskaya N, Eng KH, Khan AN, Grzankowski KS, Singel KL, Moysich K, et al. Cytokine profiling of ascites at primary surgery identifies an interaction of tumor necrosis factor-α and interleukin-6 in predicting reduced progression-free survival in epithelial ovarian cancer. Gynecol Oncol. 2015 Aug;138(2):352–7.
58. Matte I, Lane D, Laplante C, Rancourt C, Piché A. Profiling of cytokines in human epithelial ovarian cancer ascites. Am J Cancer Res. 2012;2(5):566–80.
59. Giuntoli RL 2nd, Webb TJ, Zoso A, Rogers O, Diaz-Montes TP, Bristow RE, et al. Ovarian cancer-associated ascites demonstrates altered immune environment: implications for antitumor immunity. Anticancer Res. 2009 Aug;29(8):2875–84.
60. Neill RR, Lebreton C, Bodet D, Chaibi A, Babin G, Bessede A, et al. 883 Ovarian cancer ascites exhibit an impaired immune environment – immunosuppressive & protumoral – detrimental to the immune response. J Immunother Cancer. 2024;12.
61. Amer H, Flanagan KL, Kampan NC, Itsiopoulos C, Scott CL, Kartikasari AER, et al. Interleukin-6 Is a Crucial Factor in Shaping the Inflammatory Tumor Microenvironment in Ovarian Cancer and Determining Its Hot or Cold Nature with Diagnostic and Prognostic Utilities. Cancers (Basel). 2025 May 17;17(10):1691.
62. Amer H, Kartikasari AER, Plebanski M. Elevated Interleukin-6 Levels in the Circulation and Peritoneal Fluid of Patients with Ovarian Cancer as a Potential Diagnostic Biomarker: A Systematic Review and Meta-Analysis. J Pers Med. 2021 Dec 9;11(12):1335.
63. Tang YJ, Sun ZL, Wu WG, Xing J, He YF, Xin DM, et al. Inhibitor of signal transducer and activator of transcription 3 (STAT3) suppresses ovarian cancer growth, migration and invasion and enhances the effect of cisplatin in vitro. Genet Mol Res. 2015 Mar 30;14(1):2450–60.
64. Warszyńska M, Repetowski P, Dąbrowski JM. Photodynamic therapy combined with immunotherapy: Recent advances and future research directions. Coord Chem Rev. 2023 Nov 15;495:215350.
65. Yuan Z, Fan G, Wu H, Liu C, Zhan Y, Qiu Y, et al. Photodynamic therapy synergizes with PD-L1 checkpoint blockade for immunotherapy of CRC by multifunctional nanoparticles. Mol Ther. 2021 Oct 6;29(10):2931–48.
66. Agostinis P, Berg K, Cengel KA, Foster TH, Girotti AW, Gollnick SO, et al. Photodynamic therapy of cancer: an update. CA Cancer J Clin. 2011 Jul-Aug;61(4):250-81.