Choreography and Communication
Snail embryos are textbook exemplars of autonomous cellular decision-making: in their spiral cleavage nearly every early cell division is asymmetric and oriented, creating a stereotyped and elaborate early pattern of fate-restricted cells (Figure 1A) reminiscent of the preformationist ‘homunculus’ fantasy. But the morphologically well-defined framework of these couple-dozen cells might set the stage for an array of reliably targeted, discretely detectable short-range signals. Embryos of the mud snail Ilyanassa (and thousands of other neogastropod species) make an outstanding model system for in situ single-cell analysis. It now appears that these embryos indeed have much intercellular communication to analyze.
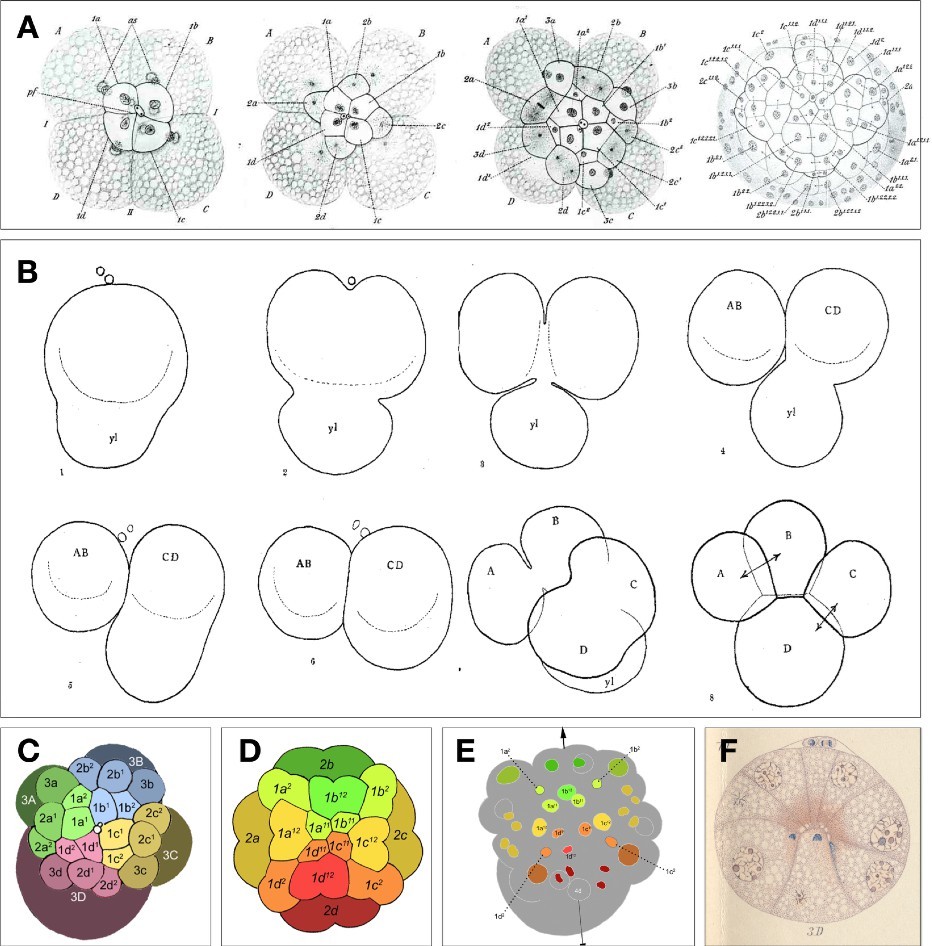
Figure 1. Early snail embryo portraits. (A) Formation of the three micromere quartets, and later ectodermal cell pattern, as shown by camera lucida drawings from The Embryology of Crepidula by Edwin G. Conklin (1897). (B) Polar lobe formation and cytoplasmic segregation to the D quadrant founder cell in Ilyanassa, as shown by camera lucida drawings of Crampton [2]; ‘yl’ stands for yolk lobe, an early moniker for the polar lobe. (C) Tracing of Craig and Morrill’s scanning electron micrograph of the 24-cell (early organizer-signaling stage) Ilyanassa embryo, with quadrant lineages color-coded and micromere tiers marked by shade. (D) Semi- diagramatic outline with axial color-coding to reflect organizer patterning of the spiralian blastula secondary axis. (E) Axially color-coded tracings of cell nuclei in Ilyanassa at a stage shortly after organizer signaling (35 cells); arrowheaded line shows the secondary body axis. Drawings in panels C, D, and E are reprinted from PLOS One [25]. (F) Watercolor camera lucida drawing of a longitudinal (frontal) section through the embryo of Physa, showing localization of distinct dye-reactive materials in basal (inward) protrusions of both micromeres and macromeres; the 3D cell is centrally internalized and in contact with apical 1q daughters as well as other cells (Wierzejski’s Plate X, Figure 10) [7].
New Clues, Unexpected Complexity
Our recently published study, employing just one simple set of microsurgical experiments in Ilyanassa [1], yielded clues that open several avenues of inquiry into pattern-forming signaling in this system. The headline item (announced in the title) is a demonstration that the famous, massive polar lobe, spectacularly shunted to the D lineage founder cell at second cleavage, is absolutely not sufficient to endow the lineage with its unique superpower of birthing a great-granddaughter cell (3D) which acts as the embryonic organizer. Very surprisingly, intercellular signaling is evidently also required. A second surprise is that this extrinsic cue seems to necessarily come from several cells in chorus, acting additively with a threshold all-or-nothing effect. A third is the identity of one of these signaling cells: the immediately adjacent (also closely related) second-quartet micromere 2d! Another surprise: two characteristic behaviors of the 3D cell seem differentially dependent on extrinsic cues, in a manner that suggests response to two separate signals from the same set of sources. Finally, not least surprising was a stark difference (between different mothers’ broods) found in the threshold requirement for 3D- inducing cell numbers; we suggested that this variability accounts for previous workers’ failure to find the extrinsic signal-dependence of 3D activation.
Two more surprising findings were off-topic. First, an enigmatic, echo-like response to 3D organizer activity (ERK activation in micromeres) was serendipitously uncoupled from 3D ERK activation; a candidate solution for this conundrum had already been published and now awaits testing. Second, we may have stumbled across an early signal in the pathway toward biomineralization.
Far from being isolated mosaic-tiles in a self-budding homunculus, then, snail blastomeres appear to engage in dynamic ritual conversation, with complex parallel signals and responses, whose intensity and/or sensitivity may be interestingly variable. But there’s more: the spiral cleavage pattern and its associated dialogue have subtly changed over hundreds of millions of years, and our story also importantly involves a macroevolutionary scenario. As explained below, an ancestral intercellular signal was previously thought to have been lost in the lineage of Ilyanassa; our study showed that it hadn’t been lost, only hidden. To appreciate our new view, some layers of background information are needed.
Background: Origin of the Ilyanassa Organizer
Our report’s highlight was evidence for one surprising signaling event, targeting a cell whose story features in most Developmental Biology textbooks as a classic example of cell autonomy. Briefly recounting this story: in one of the first embryo vivisections in history, Henry Crampton cut off the Ilyanassa zygote’s polar lobe (Figure 1B) during first cleavage before it could retract into the mother of the D quadrant founder cell; this amputation resulted in no cellular morbidity but a reliable loss of later cell diversity [2]. Experiments done half a century later identified one descendant cell of the polar lobe-inheriting D-quadrant (the 3D macromere) as the singular source of a signal that induces ectodermal pattern along the embryo's secondary axis [3,4] (Figures 1C–1E). The organizer signal’s physical nature is still uncharacterized, but its production depends on transient activation of ERK-1/2 MAP Kinase in 3D, manifested over precisely the same time window as intercellular signal transmission (deduced from timed 3D ablations) [5,6].
The textbook-upsetting main conclusion of Wandelt et al. concerns the mechanism whereby 3D becomes empowered to act as the embryonic organizer in Ilyanassa. Polar lobe inheritance is necessary but not sufficient: some intercellular signaling is additionally involved.
Deep Background: Inferences of Ancestry and Innovation
The full import of this conclusion can be appreciated only in an evolutionary context, so we have to rewind again and zoom out. The mid-20th century characterization of the Ilyanassa organizer and its apparently intrinsic determination by polar lobe inheritance was followed by extensive and incisive studies (most notably from the long-running Utrecht snail group) of distantly related gastropods whose embryos have no polar lobe, nor any asymmetry along the presumptive secondary axis. In such ‘equal-cleaving’ forms, the first two cell divisions yield four quadrant founder cells (A, B, C, D) that are identical in looks and potential.
This symmetry is broken by a dramatic intercellular signaling event. As noted by Anton Wierzejski in 1905, production of the third micromere quartet is followed by “a lively exchange… between the parts of the embryo.” [ein reger Verkehr zwischen den Teilen des Keimes… (p.534)] The four macromeres gathered around the vegetal pole (still indistinguishable) abruptly all stretch inward through the blastocoele to make contact with basal protrusions of cells at the opposite (animal) pole; invariably three of the macromeres then retract, while one remains internalized with persistent intercellular contacts: this cell becomes 3D [7] (Figure 1F).
The 3D-kissing animal pole cells, daughters of the first micromere quartet (collectively abbreviated as ‘1q’), were already known in 1905 to be the head ectoderm precursors. Now we know that, long before producing a brain and special sense organs, this sensitive and commanding set of cells plays the crucial role of ensuring that one and only one macromere is specified as organizer: when contact with 1q is prevented, no macromere becomes 3D and the embryo is left organizerless. The conditional mechanism of 3D specification (requiring 1q contact) evidently operates in embryos of the most basal members of the snail family tree [8] and in other very distantly related snails [9,10], as well as polyplacophoran mollusks [11], strongly suggesting widespread conservation of a symmetry-breaking mechanism that is at least as ancient as the phylum Mollusca [12].
Preemptive D quadrant specification by polar lobe inheritance was therefore considered an evolutionary innovation, originating just once among gastropods at the base of the very large clade (Caenogastropoda) that produced Ilyanassa. Thanks to some unknown cytoplasmic factor front-loaded by the polar lobe, the 3D cell was thought to bypass requirement for the ancestral activating signal. This thinking was bolstered by Hyla Sweet’s finding that Ilyanassa embryos deprived of all 1q cells developed as headless larvae with other organizer-dependent pattern unperturbed [13]. The same results had been obtained in Utrecht with another caenogastropod, Bithynia, whose polar lobe was known to be required for 3D identity [14]. More recently however, a surprise came from another well-studied caenogastropod, Crepidula. In these embryos, the polar lobe (much smaller than the one in Ilyanassa) is partitioned into the D quadrant founder as usual, but its experimental removal does not impair the development of organizer function [15]. The latter is instead sensitive to 1q ablation, as in primitive equal-cleaving forms [16]. An intriguing possibility thus arose: maybe the transition from conditional to autonomous 3D specification has happened gradually in caenogastropod lineages, and its steps could be deconstructed?
Wandelt’s Revolutionary Result
Or - we now think - maybe it hasn’t ever completely happened. Jessica Wandelt, studying Ilyanassa for her doctoral thesis, took an extreme approach by ablating multiple micromeres in several combinations, challenging long-held belief by asking if 3D would persistently behave like 3D. The hunch of doubt was robustly confirmed in two different experimental situations, unanimously judged by four mutually independent and qualitatively different assays. The first set of experiments gave results supporting two separately surprising conclusions. A strong immediate response was found in isolated D-quadrant quarter-embryos: after ablation of the sole 1q cell (1d), Wandelt observed total failure of ERK-1/2 ('ERK') activation in the 3D cell of most (75%) of a large sample of partial embryos. Therefore, organizer activation in Ilyanassa is not cell-autonomous. Also surprising: when both the first and second micromeres (1d and 2d) were removed from D quarter-embryos (a smaller sample of 39), 3D ERK activation was abolished in every case [1]. The strong implication of signaling from a second-quartet micromere (here, 2d) in 3D induction is unprecedented in studies of mollusk embryos. However, the experimental setup is also unprecedented, and it will be interesting to see it applied to other taxa.
Wandelt next performed multiple ablations in otherwise intact embryos, targeting combinations of micromeres 1d, 2d, and the entire first quartet, again assaying effects on 3D ERK activation. Robust effects on 3D ERK in different conditions (apparently an all-or-nothing event, probabilistically related to the number of cells removed) roughly correlated with the frequency of ectodermal organogenesis defects in identically operated embryos that were allowed to develop (~1 week) to the feeding veliger larva stage. Again, in every situation, absence of the 2d cell substantially lowered the chance of 3D ERK activation [1].
These results indicate that organizer activation is sparked by the combination of two factors: one derived intrinsically from the polar lobe and another extrinsically by signaling from multiple micromeres including 2d and several (possibly all four) 1q cells. The signaling sources seem to act in a simply additive fashion (compare Wandelt et al. Figure 2 panels D', E', and F'), suggesting that they may be delivering the same molecule, with a combined dosage that reliably crosses a definite response threshold.
How We Had All Got It Wrong
Despite the simplicity of these experiments and the compelling clarity of their results, our study also shows some sloppiness: not sloppy science but sloppy Nature, an extreme example of the nonrandom variability inherent in living systems. While assaying organogenesis following 1q ablation, postdoc Ayaki Nakamoto noted a marked inconsistency of results between different broods of embryos (of different parentage). Such variability would have delighted Darwin, and we should all be curious about it. But in this particular study's context, the documented variation is especially revealing because of Sweet's prior results with ablation of the whole first quartet: all headless veligers, otherwise normal [12], leading to the wrong conclusion. Nakamoto, performing 1q ablation on a much larger sample, obtained the same result as Sweet in a great majority of embryos (13/14) from one brood, and in small minorities (as small as 0/12) in other broods [1]. Caveat, Embryologist!
Uncoupled Responses Suggest Parallel Signaling
Three more results in the study of Wandelt et al. yield questions as much as answers. One of these is a quantitative assay of another immediate effect of micromere absence on 3D cell behavior. Normally, 3D enters mitosis and divides much earlier than its smaller, non-organizer macromere cousins 3A, 3B, and 3C. Like ERK activation, fast 3D cell cycle progression was shown to depend on the presence of multiple micromeres [1]. The dose-response curve, however, was entirely different: while ERK activation appeared to be all-or-nothing, becoming increasingly improbable with removal of more micromeres, the same increasingly severe operations resulted in progressively longer 3D cell cycles. We speculated that cell cycle progression may reflect the graded action (proportionate to combined intercellular signal strength) of an intermediate second-messenger, upstream of the all/none ERK activation [1]. However, these two neighbor-dependent behaviors of 3D are obviously not correlated: see Wandelt et al. Figure 1. A clear lack of correlation also exists between 3D cell cycle length and the ectoderm-patterning power of the organizer: compare Wandelt et al. Figure 2D and Figure 4E. While these decorrelations could artefactually reflect the known interbrood variance in signal strength and/or 3D sensitivity, one might reasonably suspect that 3D (and/or its progenitor cell(s)) may normally be receiving two separate signals concurrently.
Evidence also for Parallel Organizer Signals
Another question-yielding result concerns an oddity of Ilyanassa which stands as an evo-devo enigma: this is the secondary appearance of active ERK in a spreading crescent of all (and exclusively) the ectoderm progenitors known to receive the 3D organizer signal. The wave of ectoblast ERK activation begins while the interval of 3D ERK activation is ending, spreading laterally over the next two hours. Importantly, the wave fails to appear in embryos whose polar lobe was removed; moreover, pharmacologically blocking the wave's appearance leads to the same ectodermal organogenesis defects as polar lobe removal [5]. Oddly, the ectoblast ERK wave does not appear at all in Crepidula or other snail embryos examined so far [17–20]. In this study, it was very odd that ablation of either 1q or 2d never prevented the wave’s appearance in the remaining ectoblast, while co-ablation of 1q and 2d abolished it in every case [1]. These results too are highly decorrelated with the effects of the same ablations on 3D ERK activation as well as observed veliger-stage developmental outcomes, so we are again puzzled: we concluded that the wave is "necessary but [now we see] not sufficient" for organizer-induced development. In other words, 3D must deliver two coincident but qualitatively distinct signals to pattern the ectoderm; another implication is that both of these organizer signals depend on earlier signaling from micromeres, but in different ways. One of the ectoderm-patterning organizer signals may be DPP/BMP-2/4, which was recently shown to mediate organizer signaling in Ilyanassa [21] and a basal gastropod [22] (but oddly again, not in Crepidula) [23].
Worse than the Worst Case: Evidence for Another Signaling Target
The further we probe, indeed, the more intercellular signaling complexity emerges. The third question-yielder reported by Wandelt et al. is the extremely defective veliger-stage “Type III monster" phenotype frequently resulting from 1q ablation and almost invariably from co-ablation of 1q and 2d. With a week of incubation, organizer-deficient embryos developing after polar lobe ablation never have a shell but almost invariably display disorganized clusters of "internal shell," made of the normal aragonite [24] but deposited in abnormal places [3–6]. Type III monsters apparently don't even have that [1]. We speculated that micromeres may normally be signaling not only to 3D but to some other recipient(s) as well; an obscure old study [25] suggested a likely cellular target.
Plenty Questions…
From a snail embryo watcher’s perspective, it is no exaggeration to call this piece of work historic. The study bookends a century of inquiry, reaching back to the beginning with a gentle correction, and pointing brightly to the future with well-defined questions to be addressed by subcellular-level analyses. Proteomic (and phosphoproteomic) prospecting in dissociated and sorted single cells is an obvious next step. Another obvious approach is live imaging with fluorescent reporters: you could now, for example, quantitatively assay ERK activity in the 3D cell and subsequently in the organizer-responding ectoderm, concurrently monitoring cell division patterns, before rearing the same unique embryo to veliger stage [26]. You could target optogenetic manipulations to single cells, neighbor groups, or clones, all of which are identifiable at least as far as the 84-cell stage [27] and probably beyond [7]. Now more than ever, embryos of the mud snail and its relatives, long appreciated for their accessibility and regularity, promise to reveal unforeseeable dimensions of evolving multicellular self-organization.
Acknowledgements
The author is grateful to Lisa Nagy for extending the invitation to write this Commentary, as well as collaborating on the study to which it pertains. Thanks are due further to Jessica Wandelt and Ayaki Nakamoto, the latter now studying snail embryos across the big pond, for doing the difficult work. Across the other pond, Jo van den Biggelaar also deserves special acknowledgement for pioneering the experimental analysis of inductive D quadrant specification, and thanks for teaching us embryology at Friday Harbor Labs. Thanks of course to Gary Freeman too.
References
2. Crampton HE, Wilson EB. Experimental studies on gastropod development. Archiv für Entwicklungsmechanik der Organismen. 1896 Mar 28;3(1):1–19.
3. Clement AC. Experimental studies on germinal localization in Ilyanassa. I. The role of the polar lobe in determination of the cleavage pattern and its influence in later development. J Exp Zool. 1952 Dec;121(3):593–625.
4. Clement AC. Development of Ilyanassa following removal of the D macromere at successive cleavage stages. J Exp Zool. 1962 Apr;149(3):193–215.
5. Lambert JD, Nagy LM. MAPK signaling by the D quadrant embryonic organizer of the mollusc Ilyanassa obsoleta. Development. 2001 Jan;128(1):45–56.
6. Labordus V, Van Der Wal UP. The determination of the shell field cells during the first hour in the sixth cleavage cycle of eggs of Ilyanassa obsoleta. Journal of Experimental Zoology. 1986 Jul;239(1):65–75.
7. Wierzejski A. Embryologie von Physa fontinalis L. 1905. Leipzig: Wilhelm Engelmann. https://doi.org/10.5962/bhl.title.51967.
8. van den Biggelaar JA, Guerrier P. Dorsoventral polarity and mesentoblast determination as concomitant results of cellular interactions in the mollusk Patella vulgata. Dev Biol. 1979 Feb;68(2):462–71.
9. Martindale MQ, Doe CQ, Morrill JB. The role of animal-vegetal interaction with respect to the determination of dorsoventral polarity in the equal-cleaving spiralian, Lymnaea palustris. Wilhelm Roux' Archiv. 1985 May;194(5):281–95.
10. Boring L. Cell-cell interactions determine the dorsoventral axis in embryos of an equally cleaving opisthobranch mollusc. Dev Biol. 1989 Nov;136(1):239–53.
11. van den Biggelaar JA. Cleavage pattern and mesentoblast formation in Acanthochiton crinitus (Polyplacophora, Mollusca). Dev Biol. 1996 Mar 15;174(2):423–30.
12. Freeman G, Lundelius JW. Evolutionary implications of the mode of D quadrant specification in coelomates with spiral cleavage. J Evol Biol. 1992 Mar;5(2):205–47.
13. Sweet HC. Specification of first quartet micromeres in Ilyanassa involves inherited factors and position with respect to the inducing D macromere. Development. 1998 Oct;125(20):4033–44.
14. van Dam WI, Verdonk NH. The morphogenetic significance of the first quartet micromeres for the development of the snail Bithynia tentaculata. Wilehm Roux Arch Dev Biol. 1982 Mar;191(2):112–8.
15. Henry JQ, Perry KJ, Martindale MQ. Cell specification and the role of the polar lobe in the gastropod mollusc Crepidula fornicata. Dev Biol. 2006 Sep 15;297(2):295–307.
16. Henry JQ, Lyons DC, Perry KJ, Osborne CC. Establishment and activity of the D quadrant organizer in the marine gastropod Crepidula fornicata. Dev Biol. 2017 Nov 15;431(2):282–96.
17. Henry JJ, Perry KJ. MAPK activation and the specification of the D quadrant in the gastropod mollusc, Crepidula fornicata. Dev Biol. 2008 Jan 1;313(1):181–95.
18. Lartillot N, Lespinet O, Vervoort M, Adoutte A. Expression pattern of Brachyury in the mollusc Patella vulgata suggests a conserved role in the establishment of the AP axis in Bilateria. Development. 2002 Mar;129(6):1411–21.
19. Lambert JD, Nagy LM. The MAPK cascade in equally cleaving spiralian embryos. Dev Biol. 2003 Nov 15;263(2):231–41.
20. Koop D, Richards GS, Wanninger A, Gunter HM, Degnan BM. The role of MAPK signaling in patterning and establishing axial symmetry in the gastropod Haliotis asinina. Dev Biol. 2007 Nov 1;311(1):200–12.
21. Lambert JD, Johnson AB, Hudson CN, Chan A. Dpp/BMP2-4 Mediates Signaling from the D-Quadrant Organizer in a Spiralian Embryo. Curr Biol. 2016 Aug 8;26(15):2003–10.
22. Tan S, Huan P, Liu B. Molluskan Dorsal-Ventral Patterning Relying on BMP2/4 and Chordin Provides Insights into Spiralian Development and Evolution. Mol Biol Evol. 2022 Jan 7;39(1):msab322.
23. Lyons DC, Perry KJ, Batzel G, Henry JQ. BMP signaling plays a role in anterior-neural/head development, but not organizer activity, in the gastropod Crepidula fornicata. Dev Biol. 2020 Jul 15;463(2):135–57.
24. McCain ER. Cell interactions influence the pattern of biomineralization in the Ilyanassa obsoleta (Mollusca) embryo. Dev Dyn. 1992 Nov;195(3):188–200.
25. Cather JN. Cellular interactions in the development of the shell gland of the gastropod, Ilyanassa. J Exp Zool. 1967 Nov;166(2):205–23.
26. Mandell JW, Gocan NC. A green fluorescent protein kinase substrate allowing detection and localization of intracellular ERK/MAP kinase activity. Anal Biochem. 2001 Jun 15;293(2):264–8.
27. Goulding MQ. Cell lineage of the Ilyanassa embryo: evolutionary acceleration of regional differentiation during early development. PLoS One. 2009;4(5):e5506.