Abstract
To maintain identity and epigenetic balance, a cell relies on robust transcriptional programs regulated both by transcription factors and by dynamic chromatin states. These states, which enable or restrict gene expression and genome function, are shaped by the incorporation of histone variants into nucleosomes and their post-translational modifications. While canonical histones, such as H3.1, and variants like H3.3 are well-characterized and conserved across species, other histone variants exhibit organism-specific functions and remain largely unexplored. A recent study [1] identifies H3.14 as a novel Arabidopsis thaliana histone H3 variant that is rapidly and transiently induced upon abiotic stress in a defined subset of epidermal cells in the root transition zone. Loss of H3.14 leads to increased root growth under salt stress and an enlargement of the transition zone, while the root apical meristem remains unaffected. Chromatin immunoprecipitation and transcriptomic data reveal that H3.14 exhibits distinct deposition patterns associated with the activation of stress response genes and the repression of growth-related genes. Here, we discuss the implications of these findings, present additional evidence consistent with a post-transcriptional mechanism controlling H3.14 spatial specificity and integrate H3.14 dynamics into a working temporal model of the root stress response.
Keywords
Chromatin, Epigenetics, Histone Variant, Salt stress, Drought, Arabidopsis, Plant
H3.14 is Induced by Abiotic Stress and Has a Dual Transcriptional Role
The histone H3 gene family in Arabidopsis thaliana is composed of 7 members encoded by 15 genes [2]. The HTR14 gene (At1g75600), encoding H3.14, contains upstream DNase-I-accessible regions with putative binding sites for DREB, ABRE, NAC, and WRKY transcription factors, all associated with the response to abiotic stress [3,4]. Accordingly, exposure to salt stress (NaCl), abscisic acid (ABA), and reactive oxygen species (ROS) rapidly and transiently induces HTR14 transcription. A key question is how the off/on switch of HTR14 transcription is established. Under normal conditions, the HTR14 locus is covered by the silencing mark H3K27me3, which rapidly disappears at the proximal promoter and 5' end upon salt treatment. Intriguingly, the active mark H3K4me3 is present in the locus both before and after stress, suggestive of bivalent chromatin, a configuration with both marks that keep genes poised for rapid activation [5]. As a consequence, during stress exposure, H3.14 protein rapidly accumulates in a subset of epidermal cells of the transition zone (TZ) of the root, the first organ to sense stress cues in the soil [6].
The physiological relevance of H3.14 was examined using htr14 knockdown lines. After NaCl treatment, the roots of these plants exhibited enhanced growth, whereas constitutive H3.14 expression was deleterious to root growth both in the absence and presence of NaCl. At the cellular level, the number of cortical cells in TZ decreased in wild-type plants after salt treatment but not in the htr14 mutants.
At the genomic level, H3.14 showed three deposition patterns after NaCl treatment: enrichment at both proximal promoters and transcription termination sites (TTS), associated with upregulated stress response genes (cluster 1); preferential enrichment at proximal promoters, associated with downregulated growth genes (cluster 2); and enrichment at TTS, associated with housekeeping genes with unchanged expression (cluster 3). Transcriptomics data confirmed a dual role of H3.14: 67% of genes upregulated in the mutant were downregulated in wild type under salt stress, and 57% of downregulated genes in the mutant were upregulated in wild type. H3.14 thus functions at both proximal promoters and TTSs to activate stress response genes, and at proximal promoters alone to repress growth-related genes. How these differential deposition patterns are established remains to be tested. Notably, the third pattern — enrichment at TTS of housekeeping genes — resembles the 3' enrichment described for H3.3 [7–9]. This deposition is characteristic of replacement histones, where nucleosomes displaced by RNA Pol II are restored through a gap-filling mechanism [10,11]. The rapid turnover of H3.14 observed in FRAP experiments, dependent on active transcription, supports this interpretation.
H3.14 Spatial Specificity Extends beyond Transcriptional Control
An intriguing observation reported in the article is that constitutive H3.14 expression under the UBI10 promoter (pUBI10::HTR14-GFP) is deleterious for root growth, but leaves open the question of whether the spatial restriction of H3.14 to the TZ depends solely on the HTR14 promoter. In transgenic plants expressing HTR14 under the constitutive UBI10 promoter, one would expect uniform H3.14 incorporation across all root cells. However, under normal growth conditions, while H3.14-GFP was detected across all tissue layers and root domains, the signal was not uniform. A preferential deposition was observed within the epidermal cells of the TZ, with enhanced GFP signal in this region even in the absence of stress (Figure 1). Under salt stress, this preferential deposition became even more pronounced, with a marked increase in H3.14 signal in the TZ. Quantification of GFP fluorescence intensity confirmed significantly higher levels in the TZ compared to the root apical meristem (RAM), both in control and stress conditions.
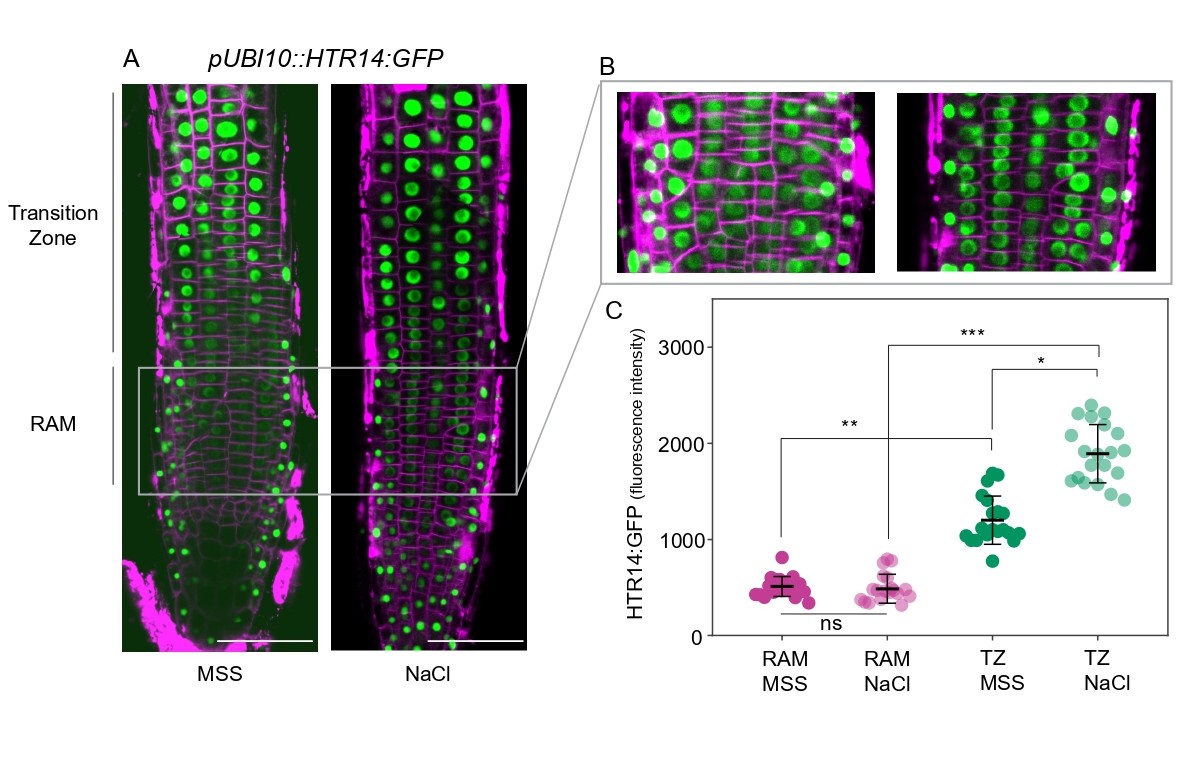
Figure 1. Preferential deposition of H3.14 in the transition zone of pUBI10::HTR14:GFP constitutive expression lines. (A) Confocal microscopy images of roots grown under normal conditions (MSS) and after 4 hours of 140 mM NaCl treatment. Magenta indicates cell membranes stained with FM4-64, and green represents H3.14-GFP. (B) Cropped images of the root apical meristem (RAM) region under MSS and NaCl conditions. (C) Quantification of H3.14-GFP fluorescence intensity in the RAM and transition zone under MSS and NaCl conditions. Error bars represent the SEM, and statistical significance was determined using one-way ANOVA followed by Tukey’s post-hoc multiple-comparison test. ***, p < 0.001; **, p < 0.01; *, p < 0.05; ns, not significant (RAM MSS vs RAM NaCl). Scale bars: 50 μm. Each data point represents fluorescence intensity of an individual epidermal nucleus, pooled from 3 independent roots per condition. All images were acquired under identical confocal settings (laser power, gain and acquisition parameters) to ensure reproducibility.
These results indicate that the spatial restriction of H3.14 to the TZ extends beyond transcriptional activation by the HTR14 promoter and that post-transcriptional and/or deposition-level mechanisms contribute as well. Several non-exclusive scenarios may explain this preferential accumulation: differential protein stability, differential nucleosome turnover, differential translation efficiency, or residual heterogeneity in UBI10 promoter activity across root cell types. To minimize the latter, quantifications in Figure 1 were restricted to epidermal nuclei. A mechanistically attractive possibility is the existence of a chaperone preferentially active in the TZ, which would recognize H3.14 based on its amino acid sequence and/or its relative abundance. Consistent with this idea, the article reports that H3.14 deposition is independent of both HIRA and CAF-1, and therefore lies outside the two canonical histone H3 assembly pathways. More broadly, factors with auxiliary chaperone activity, dispensable under homeostatic conditions, may become engaged when the chromatin assembly system is challenged by stress.
H3.14 Dynamics Correlate with the Temporal Phases of the Stress Response
The live-imaging data revealed three distinct phases of root elongation under NaCl stress: a stop phase (0–4 h), a quiescent phase (4–8 h), and a recovery phase (8–12 h), consistent with earlier reports [6]. H3.14 accumulates during the stop and quiescent phases and decreases during the recovery phase (Figure 2A). By quantifying the fluorescence intensity of H3.14-GFP relative to the constitutively expressed H3.3-mRFP in individual nuclei over time, it was possible to classify nuclei into groups based on their H3.14/H3.3 ratio (Figure 2B). Nuclei with the highest fluorescence intensity ratio maintained H3.14 for more than 10 hours and were consistently located in the TZ (Figure 2C). Nuclei with intermediate ratios (close to 2) exhibited an induction period of approximately 4–5 hours followed by decay and were distributed over a broader region of the TZ. Nuclei with a ratio close to or below 1 showed very transient H3.14 expression and were located in the RAM or elongation zone (Figure 2C). The spatial distribution of H3.14 residence time reinforces its functional specificity for the TZ.
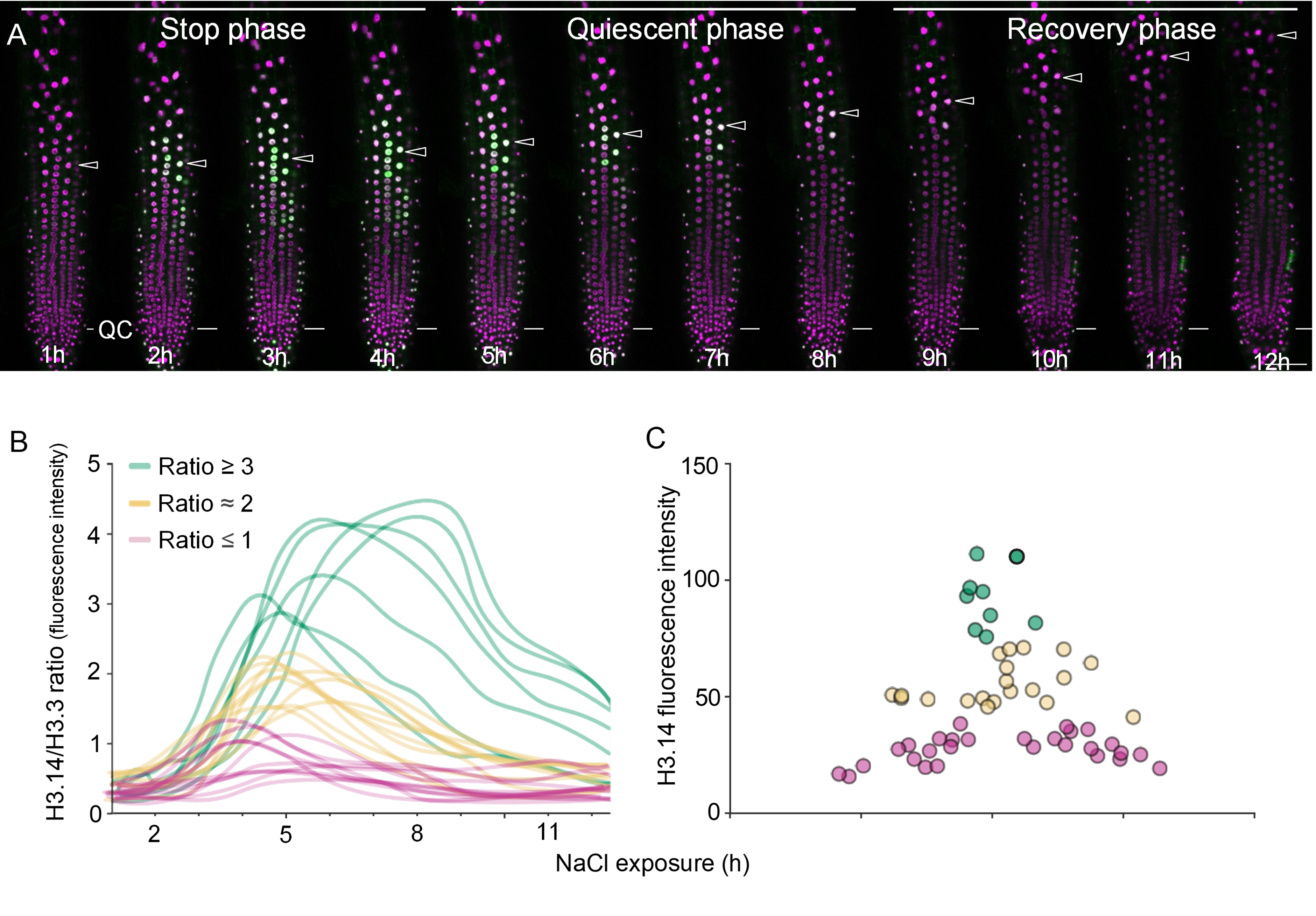
Figure 2. Dynamics of H3.14-GFP expression in response to salt stress. (A) Live-imaging of one 5-day-old root expressing H3.14-GFP (green) and H3.3-mRFP (magenta) during 12 hours treatment with 140 mM NaCl. Arrowheads mark the same nucleus across consecutive time frames. Scale bar: 50 μm. Reprinted from Figure 3D of [1], with permission from Elsevier. This is the only panel reprinted from the original article, with permission; the rest are original. (B) Temporal analysis of the H3.14/H3.3 fluorescence intensity ratio in individual nuclei during salt stress exposure. Total n= 60 nuclei from three roots. Nuclei with high H3.14/H3.3 ratios (>3) exhibit longer residence times of H3.14, while intermediate (ratio ~2) and low (ratio ~1) nuclei show shorter durations of H3.14 retention. (C) Spatial distribution of H3.14-positive nuclei along an epidermal cell file. High fluorescence intensity nuclei are primarily localized in the transition zone (position 1-2), with lower-intensity nuclei found in the root apical meristem (position 0-1) and elongation zone (position 2-3). The color of each data point corresponds to the H3.14/H3.3 fluorescence intensity ratio described in panel B.
The H3.14 dynamics revealed two relevant concepts. One is the H3.14 deposition and maintenance seems to be specifically regulated to occur in the TZ. Another is that an anti-correlation temporal link exists between root growth and the presence of H3.14. These data, integrated with the transcriptomic and ChIP-seq results, support a temporal model (Figure 3). During the stop phase, HTR14 mRNA peaks and H3.14 is deposited at stress response genes and growth-related genes. During the quiescent phase, HTR14 transcription declines but H3.14 protein remains sufficient to sustain the transcriptional program, as directly shown by the live-imaging signal in Figure 2A. As H3.14 protein levels decay around 6 hours, growth-related genes start to recover. During the recovery phase, H3.14 becomes undetectable and cells resume elongation.
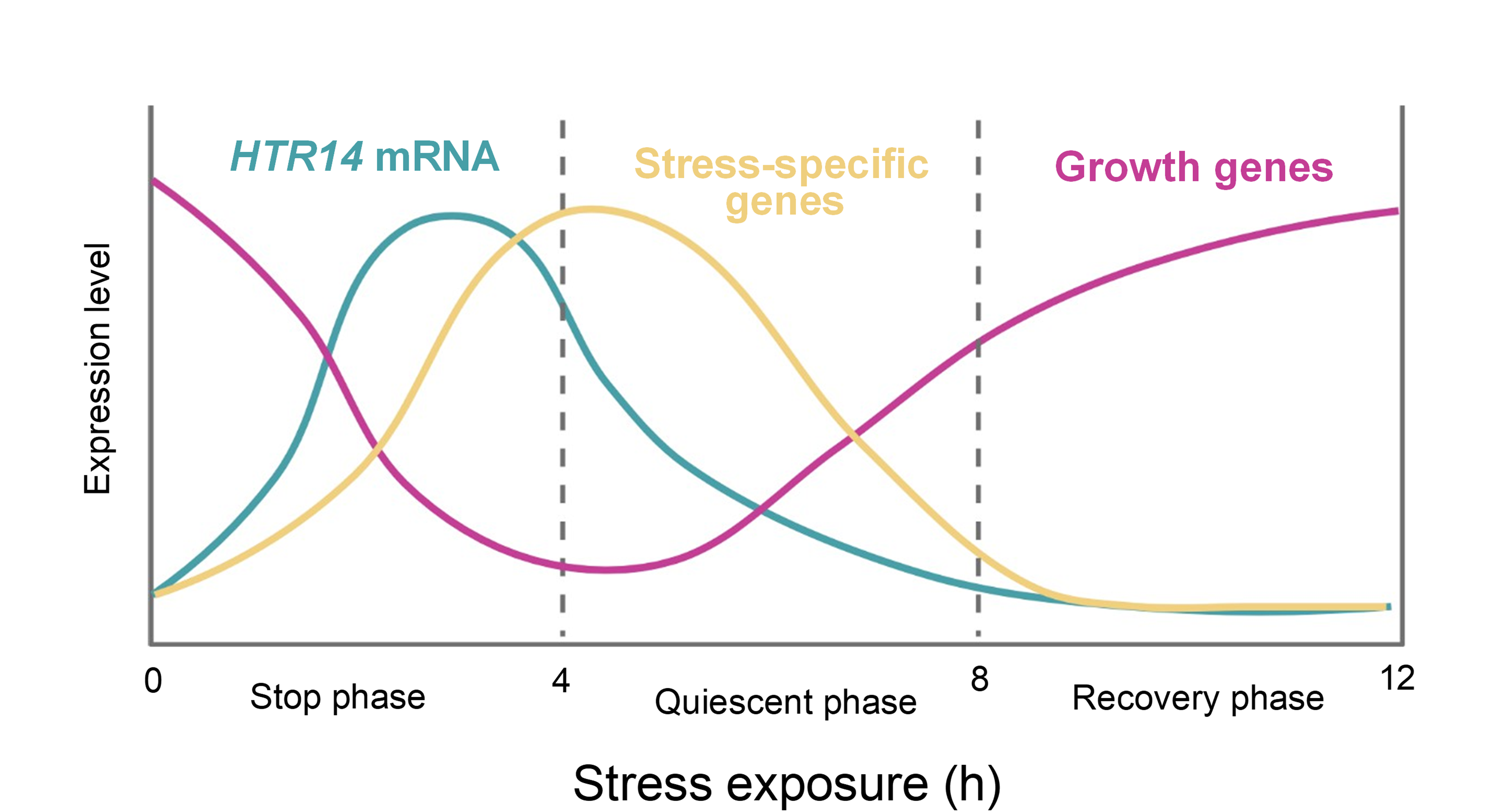
Figure 3. Working model depicting the temporal expression patterns of growth-related genes (pink), stress-specific early response genes (yellow), and HTR14 mRNA (blue) after initiation of stress exposure (time 0). The stop phase (0–4 h) is characterized by a peak in HTR14 mRNA, which precedes the activation of stress-specific genes and the repression of growth-related genes. During the quiescent phase (4–8 h), HTR14 transcription declines but H3.14 protein remains detectable in transition-zone nuclei, sustaining the transcriptional output (see protein-level dynamics in Figure 2A). In the recovery phase (8–12 h), H3.14 protein becomes undetectable and growth-related genes recover expression, allowing the resumption of elongation.
The Transition Zone as a Chromatin-regulated Developmental Boundary
The epidermal cells of the TZ are constantly exposed to the soil environment, as opposed to the meristem, which is protected by the lateral root cap [12,13]. Consistent with this, several studies revealed that TZ is more sensitive to abiotic stress than other root regions. For instance, phosphate deficiency triggers Fe³+ accumulation and ROS generation in the TZ, causing cell wall stiffening and rapid inhibition of cell elongation [14,15]. The balance of reactive oxygen species (O2·_ and H2O2) also regulates the division-to-elongation transition through the transcription factor UPB1, which directly represses peroxidase genes at this boundary [16,17]. Similarly, in the presence of aluminum, local auxin biosynthesis via YUC and TAA pathways inhibits growth specifically at the TZ [18]. Salt stress itself causes immediate growth arrest at the TZ prior to other root regions [6]. These examples converge on TZ as a hub for stress perception and growth regulation.
These findings gain additional significance when placed in a developmental context. The TZ marks the boundary where cells switch from division to elongation [19]. This transition is positioned by an auxin minimum, generated through the activity of the cytokinin-dependent transcription factor ARR1. ARR1 activates SHY2, a repressor of PIN auxin efflux carriers, and GH3.17, which conjugates active auxin [20,21]. ARR1 also activates proton-pump ATPases (AHA1, AHA2) and the expansin EXPA1, promoting cell wall acidification and loosening required for elongation [22,23]. Transcriptomics data from the htr14-1 mutant show that H3.14 negatively regulates the expression of ARR1, GH3.17, EXPA1, AHA1, and AHA2. Interestingly, ChIP-seq data show that H3.14 is deposited at the proximal promoter of these genes, matching the deposition pattern of H3.14-repressed targets. This raises the possibility that H3.14 participates in the auxin-cytokinin signaling network within the TZ during stress response.
Concluding Remarks and Perspectives
The original work discussed here [1] identified H3.14 as a histone variant dedicated to the early abiotic stress response in the root. The dual deposition pattern — at both proximal promoters and TTSs in active genes and at proximal promoters alone in repressed genes — provides a mechanism for the coordinated regulation of stress and growth genes. The additional data presented here show that H3.14 spatial distribution specificity does not rely solely on promoter activity but involves post-transcriptional mechanisms that preferentially direct its incorporation to TZ. This dynamic reveals a graded response, with the longest H3.14 residence time in TZ cells and transient expression at the periphery of its expression domain.
Several questions remain: (i) the mechanism of H3.14 deposition, independent of HIRA and CAF-1, needs to be identified, (ii) how H3.14-containing nucleosomes generate opposing transcriptional outputs depending on their genomic position is not known, (iii) whether H3.14 cooperates with other chromatin features — such as H2A.Z, whose targets significantly overlap with H3.14 targets [24], among others, remain to be tested. Furthermore, the connection between H3.14 and the auxin-cytokinin network in TZ could provide insight into how chromatin remodeling interfaces with hormone-regulated developmental boundaries.
Finally, the finding that a single histone variant can orchestrate a transcriptional response in a defined spatial-temporal manner opens new avenues for understanding how chromatin is remodeled during stress. Whether the H3.14 paradigm applies to other plant-specific histone variants or to stress-inducible variants in other organisms is also an area for future exploration. In primates, H3.X and H3.Y are induced under cellular stress and high cell density [25] and notably share amino acid substitutions with H3.14 at four of the five H3.14-specific positions, despite their independent evolution. In rice, the Oryza-specific variant H4.V is redistributed under salt stress to enable H4 lysine 5 acetylation at protein-coding loci [26]. The convergence of these examples argues that the emergence of stress-specialized variants represents a recurrent evolutionary solution to the demand for rapid, reversible transcriptional reprogramming.
Acknowledgements
Research at the C.G. laboratory is funded by PID2021-123319NB-I00 and PID2024-159682NB-I00 (Ministerio de Ciencia, Innovación y Universidades), 202620E028 (CSIC). The Centro de Biología Molecular Severo Ochoa is the recipient of a ‘Severo Ochoa Excellence’ grant (CEX2021-001154-S) funded by MICIU/AEI (10.13039/501100011033) and receives institutional support from the Ramón Areces Foundation.
References
2. Nunez-Vazquez R, Desvoyes B, Gutierrez C. Histone variants and modifications during abiotic stress response. Front Plant Sci. 2022 Dec 15;13:984702.
3. Hubbard KE, Nishimura N, Hitomi K, Getzoff ED, Schroeder JI. Early abscisic acid signal transduction mechanisms: newly discovered components and newly emerging questions. Genes Dev. 2010 Aug 15;24(16):1695–708.
4. Shah WH, Rasool A, Saleem S, Mushtaq NU, Tahir I, Hakeem KR, et al. Understanding the Integrated Pathways and Mechanisms of Transporters, Protein Kinases, and Transcription Factors in Plants under Salt Stress. Int J Genomics. 2021 Apr 12;2021:5578727.
5. Faivre L, Schubert D. Facilitating transcriptional transitions: an overview of chromatin bivalency in plants. J Exp Bot. 2023 Mar 28;74(6):1770–83.
6. Geng Y, Wu R, Wee CW, Xie F, Wei X, Chan PM, et al. A spatio-temporal understanding of growth regulation during the salt stress response in Arabidopsis. Plant Cell. 2013 Jun;25(6):2132–54.
7. Stroud H, Otero S, Desvoyes B, Ramírez-Parra E, Jacobsen SE, Gutierrez C. Genome-wide analysis of histone H3.1 and H3.3 variants in Arabidopsis thaliana. Proc Natl Acad Sci U S A. 2012 Apr 3;109(14):5370–5.
8. Wollmann H, Holec S, Alden K, Clarke ND, Jacques PÉ, Berger F. Dynamic deposition of histone variant H3.3 accompanies developmental remodeling of the Arabidopsis transcriptome. PLoS Genet. 2012;8(5):e1002658.
9. Zhao T, Lu J, Zhang H, Xue M, Pan J, Ma L, et al. Histone H3.3 deposition in seed is essential for the post-embryonic developmental competence in Arabidopsis. Nat Commun. 2022 Dec 13;13(1):7728.
10. Ray-Gallet D, Woolfe A, Vassias I, Pellentz C, Lacoste N, Puri A, et al. Dynamics of histone H3 deposition in vivo reveal a nucleosome gap-filling mechanism for H3.3 to maintain chromatin integrity. Mol Cell. 2011 Dec 23;44(6):928–41.
11. Delaney K, Mailler J, Wenda JM, Gabus C, Steiner FA. Differential Expression of Histone H3.3 Genes and Their Role in Modulating Temperature Stress Response in Caenorhabditis elegans. Genetics. 2018 Jun;209(2):551–65.
12. Berhin A, de Bellis D, Franke RB, Buono RA, Nowack MK, Nawrath C. The Root Cap Cuticle: A Cell Wall Structure for Seedling Establishment and Lateral Root Formation. Cell. 2019 Mar 7;176(6):1367–78.e8.
13. Kollist H, Zandalinas SI, Sengupta S, Nuhkat M, Kangasjärvi J, Mittler R. Rapid Responses to Abiotic Stress: Priming the Landscape for the Signal Transduction Network. Trends Plant Sci. 2019 Jan;24(1):25–37.
14. Balzergue C, Dartevelle T, Godon C, Laugier E, Meisrimler C, Teulon JM, et al. Low phosphate activates STOP1-ALMT1 to rapidly inhibit root cell elongation. Nat Commun. 2017 May 15;8:15300.
15. Mora-Macías J, Ojeda-Rivera JO, Gutiérrez-Alanís D, Yong-Villalobos L, Oropeza-Aburto A, Raya-González J, et al. Malate-dependent Fe accumulation is a critical checkpoint in the root developmental response to low phosphate. Proc Natl Acad Sci U S A. 2017 Apr 25;114(17):E3563–72.
16. Tsukagoshi H, Busch W, Benfey PN. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell. 2010 Nov 12;143(4):606–16.
17. Dunand C, Crèvecoeur M, Penel C. Distribution of superoxide and hydrogen peroxide in Arabidopsis root and their influence on root development: possible interaction with peroxidases. New Phytol. 2007;174(2):332–41.
18. Liu G, Gao S, Tian H, Wu W, Robert HS, Ding Z. Local Transcriptional Control of YUCCA Regulates Auxin Promoted Root-Growth Inhibition in Response to Aluminium Stress in Arabidopsis. PLoS Genet. 2016 Oct 7;12(10):e1006360.
19. Kong X, Liu G, Liu J, Ding Z. The Root Transition Zone: A Hot Spot for Signal Crosstalk. Trends Plant Sci. 2018 May;23(5):403–9.
20. Di Mambro R, De Ruvo M, Pacifici E, Salvi E, Sozzani R, Benfey PN, et al. Auxin minimum triggers the developmental switch from cell division to cell differentiation in the Arabidopsis root. Proc Natl Acad Sci U S A. 2017 Sep 5;114(36):E7641–9.
21. Pierdonati E, Unterholzner SJ, Salvi E, Svolacchia N, Bertolotti G, Dello Ioio R, et al. Cytokinin-Dependent Control of GH3 Group II Family Genes in the Arabidopsis Root. Plants (Basel). 2019 Apr 8;8(4):94.
22. Pacifici E, Di Mambro R, Dello Ioio R, Costantino P, Sabatini S. Acidic cell elongation drives cell differentiation in the Arabidopsis root. EMBO J. 2018 Aug 15;37(16):e99134.
23. Cosgrove DJ. Plant expansins: diversity and interactions with plant cell walls. Curr Opin Plant Biol. 2015 Jun;25:162–72.
24. Coleman-Derr D, Zilberman D. Deposition of histone variant H2A.Z within gene bodies regulates responsive genes. PLoS Genet. 2012;8(10):e1002988.
25. Wiedemann SM, Mildner SN, Bönisch C, Israel L, Maiser A, Matheisl S, et al. Identification and characterization of two novel primate-specific histone H3 variants, H3.X and H3.Y. J Cell Biol. 2010 Sep 6;190(5):777–91
26. Gandhivel VH, Sotelo-Parrilla P, Raju S, Jha S, Gireesh A, Harshith CY, et al. An Oryza-specific histone H4 variant predisposes H4 lysine 5 acetylation to modulate salt stress responses. Nat Plants. 2025 Apr;11(4):790-807.