Abstract
VQ motif-containing proteins represent a plant-specific class of transcriptional cofactors that integrate developmental and stress-responsive signaling pathways. Recent genome-wide characterization of the VQ gene family in the recretohalophyte L. bicolor identified LbVQ6 as a key regulator of salt gland development and salinity tolerance. Here, we synthesize previously published evidence on the signaling role of LbVQ6 and position it within regulatory networks involving MAPK-mediated phosphorylation cascades, WRKY transcription factor interactions, phytohormone signaling, and redox homeostasis. Functional silencing of LbVQ6 as reported in earlier studies results in a marked reduction in salt gland density, disruption of ionic balance, and increased accumulation of reactive oxygen species, demonstrating its essential role in both structural and physiological adaptation to saline environments. Promoter cis-element analysis further suggests that LbVQ6 integrates ABA-, JA-, and SA-dependent stress signaling pathways, although functional validation of these regulatory interactions remains necessary. Collectively, these findings highlight VQ-mediated regulatory networks as critical components of halophyte stress adaptation. The identification of LbVQ6-centered signaling pathways provides promising molecular targets for engineering salinity tolerance in crop species. Future studies focusing on protein interaction networks, post-translational regulation, and downstream transcriptional programs will further clarify the role of VQ proteins in plant stress signaling.
Keywords
VQ protein, Signaling pathway, Salt tolerance, Halophyte, Transcriptional regulation, Salt gland development, L. bicolor
Introduction
Global soil salinization is one of the most severe abiotic constraints limiting agricultural productivity worldwide. More than 20% of irrigated land and nearly 7% of terrestrial ecosystems are affected by salinity, resulting in osmotic imbalance, ionic toxicity, oxidative stress, and metabolic disruption in plants [1,2]. Elucidating the molecular signaling mechanisms underlying salt tolerance is therefore a major objective in plant biology and crop improvement.
Plant responses to salt stress are regulated by complex signaling networks involving calcium signaling, mitogen-activated protein kinase (MAPK) cascades, phytohormone pathways, and transcriptional regulatory systems [2,3]. Within these networks, transcriptional cofactors play key roles in linking upstream signaling with downstream gene expression.
VQ motif-containing proteins represent an important class of such cofactors, defined by the conserved FxxhVQxhTG motif that mediates interactions with WRKY transcription factors and MAP kinases [4,5]. These proteins regulate plant growth, development, and stress responses by acting as intermediates between signaling cascades and transcriptional networks [4–6]. Functional studies demonstrate that VQ proteins modulate stress adaptation through interactions with transcription factors and kinase signaling pathways [3,5,7,8].
At the molecular level, VQ proteins contribute to stress tolerance by regulating ion transport, reactive oxygen species (ROS) detoxification, and osmotic balance [2,12], thereby enabling coordinated physiological and transcriptional responses to environmental stress.
Halophytes provide valuable models for studying salt tolerance due to their ability to thrive under high salinity. These plants possess specialized adaptations such as ion compartmentalization, selective ion transport, and salt secretion systems [1,13]. In recretohalophytes, salt glands actively remove excess ions, maintaining cellular ion homeostasis [13].
L. bicolor is a representative recretohalophyte with multicellular salt glands that are critical for salt tolerance. Recent studies have identified key regulatory genes involved in salt gland development, including members of the VQ gene family [6,13]. Genome-wide analyses of VQ gene families in different plant species, including wheat, bamboo, and poplar, have revealed diverse gene numbers and functional roles in stress signaling [9–11]. Among them, LbVQ6 functions as a positive regulator of salt gland development and salt tolerance. Silencing of LbVQ6 results in reduced salt gland density, increased ROS accumulation, and disrupted ion balance, highlighting its role in coordinating developmental and stress-response pathways.
Despite these advances, the signaling mechanisms by which VQ proteins regulate stress adaptation in halophytes remain unclear. In particular, how VQ-mediated pathways integrate developmental, hormonal, and environmental signals represents a key unresolved question. Addressing this gap will advance our understanding of plant stress tolerance and support the development of stress-resilient crops.
This article is presented as a commentary synthesizing previously published experimental and genomic data, primarily derived from studies such as Yang [6] and Yuan [14]. The objective is to critically evaluate existing evidence and propose integrative signaling models explaining the role of LbVQ6 in coordinating developmental and stress-responsive pathways in halophytes.
Evolutionary Significance of VQ Gene Family Contraction in Halophytes
Genome-wide comparative analyses of VQ motif-containing gene families across plant species have revealed substantial variation in gene copy number, reflecting lineage-specific evolutionary dynamics and potential functional diversification. In glycophytic model plants such as A. thaliana, 34 VQ genes have been identified, whereas larger gene families have been reported in crops such as soybean (G. max, 74 members), rice (O. sativa, 39 members), and tomato (S. lycopersicum, 26 members) [4–6,14]. In contrast, genome-wide analysis of the halophyte L. bicolor revealed the presence of only six VQ genes, representing a marked reduction compared to glycophytic species. This difference suggests a possible contraction of the VQ gene family; however, its adaptive significance requires careful interpretation.
Gene family contraction is a recognized evolutionary process that can arise through selective pressures eliminating redundant paralogous genes while retaining essential regulatory components [15,16]. In specialized ecological niches such as saline environments, genome streamlining has been proposed as a mechanism that may enhance regulatory efficiency by reducing redundancy and simplifying signaling networks [17,58]. From this perspective, the reduced number of VQ genes in L. bicolor could reflect selective retention of core regulatory elements required for stress signaling and salt gland development.
However, this interpretation should be treated cautiously. Gene family contraction may also result from alternative processes, including lineage-specific genome evolution, incomplete genome assembly, or annotation limitations rather than direct adaptive selection [18]. Furthermore, apparent reduction in gene number does not necessarily imply reduced functional complexity, as compensatory mechanisms involving other transcription factor families or signaling pathways may exist. Therefore, the observed contraction of the VQ gene family in L. bicolor should not be considered definitive evidence of adaptive optimization without broader comparative and functional validation.
Phylogenetic analysis, as reported in previous studies [6], indicates that the six LbVQ proteins are distributed among four subfamilies (II, III, IV, and V), while subfamily I-commonly present in glycophytic species-is absent in L. bicolor. This subgroup-specific loss may indicate that certain regulatory pathways mediated by subgroup I VQ proteins are either dispensable in halophytes or functionally replaced by alternative regulatory mechanisms. Functional redundancy among paralogous genes often permits gene loss without compromising biological function, particularly when compensatory pathways are present [6].
Gene family contraction has been associated with adaptation to extreme environments in several plant systems, where reduced gene numbers may coincide with increased functional specialization [15,17]. In halophytes, specialized structures such as salt glands provide highly efficient mechanisms for ion secretion and homeostasis, potentially reducing the requirement for extensive regulatory redundancy [13]. Nevertheless, it remains unclear whether reduced VQ gene number directly contributes to stress adaptation or simply reflects broader genome evolutionary patterns.
Another notable feature of LbVQ genes is their predominantly intronless structure, which has also been reported in other plant species, including A. thaliana, rice, and tomato [4–6,14]. Intron loss has sometimes been associated with rapid transcriptional responses; however, this relationship is not universal. Introns can also enhance gene expression, facilitate alternative splicing, and contribute to regulatory complexity [19]. Therefore, the functional implications of intron loss in LbVQ genes remain uncertain and require experimental validation rather than being interpreted solely as an adaptation for rapid expression.
Comparative genomic analysis further revealed conserved collinearity relationships between L. bicolor and other plant species, suggesting that the retained LbVQ genes have been evolutionarily conserved due to essential biological roles. Functional studies have demonstrated that LbVQ6 contributes to salt gland development and salt tolerance, indicating that at least some retained VQ genes play critical regulatory roles in halophyte adaptation [6].
Taken together, the apparent contraction of the VQ gene family in L. bicolor may reflect a combination of evolutionary processes, including selective retention, genome streamlining, and lineage-specific genomic variation. While this pattern could be associated with functional optimization under saline conditions, definitive conclusions require further comparative genomic studies across multiple halophytes and direct functional analyses. These findings highlight both the potential significance and the current uncertainty surrounding VQ gene evolution in plant stress signaling networks (Table 1).
|
Species |
Number of VQ genes |
Environmental adaptation |
Reference |
|
Arabidopsis thaliana |
34 |
Glycophyte |
[4] |
|
Glycine max |
74 |
Glycophyte |
[10] |
|
Oryza sativa |
39 |
Glycophyte |
[55] |
|
Solanum lycopersicum |
26 |
Glycophyte |
[66] |
|
Limonium bicolor |
6 |
Halophyte |
[6] |
LbVQ6 as a Central Signaling Hub in Salt Gland Development
Salt glands are specialized epidermal structures responsible for salt secretion in recretohalophytes and represent a key adaptive mechanism enabling plants to survive under high salinity conditions. These structures actively transport and secrete excess sodium ions, thereby maintaining cellular ion homeostasis and preventing ion toxicity [13]. In L. bicolor, salt glands are multicellular and highly organized, and their density and functional activity are closely associated with salt tolerance capacity. Their development requires coordinated regulation of epidermal cell differentiation and transcriptional signaling networks [20]. Based on available evidence, LbVQ6 is proposed to function within a broader signaling network regulating salt gland development and stress adaptation (Figure 1).
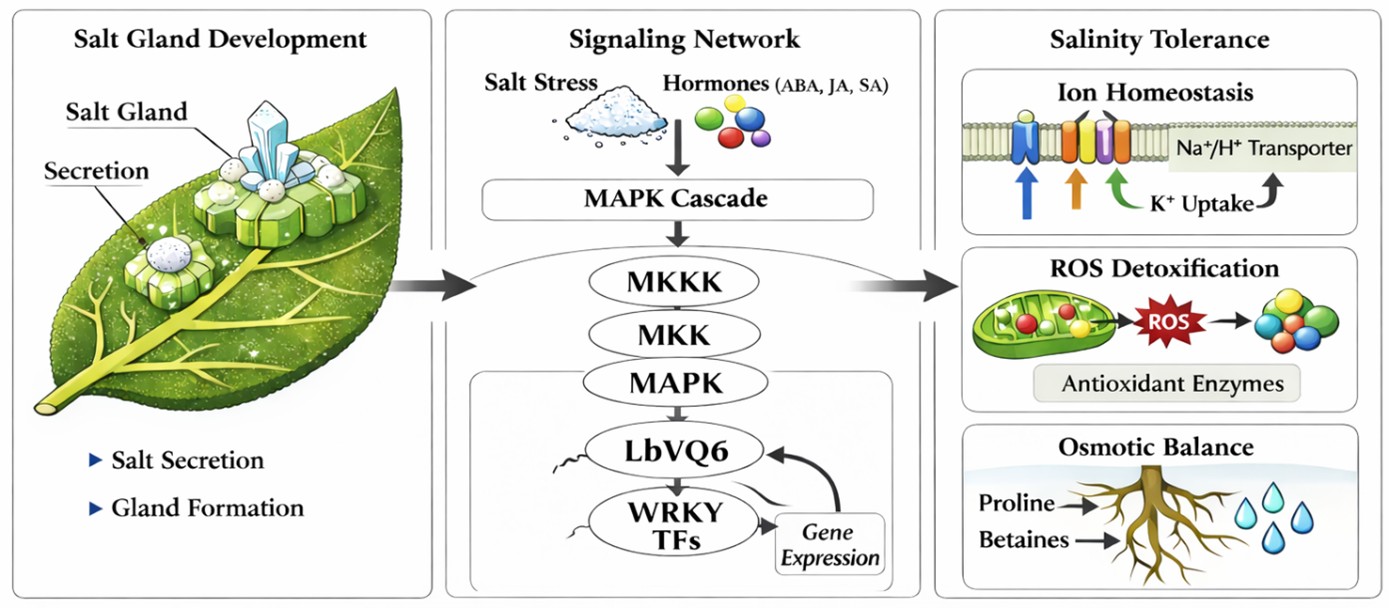
Figure 1. Proposed signaling network of LbVQ6 in regulating salt gland development and salt stress tolerance in Limonium bicolor.
Experimental evidence supporting the role of LbVQ6 in salt gland development is primarily derived from virus-induced gene silencing (VIGS) studies. Silencing of LbVQ6 resulted in a significant reduction in salt gland density, accompanied by decreased salt secretion capacity and increased sensitivity to salt stress [6]. In addition to morphological effects, LbVQ6 silencing was associated with increased accumulation of reactive oxygen species and disruption of ion homeostasis [12]. These findings provide direct evidence that LbVQ6 contributes to both developmental and physiological processes required for salt tolerance.
At the transcriptional level, LbVQ6 silencing was shown to alter the expression of genes associated with epidermal cell differentiation, including LbEGL3 (positive regulator) and LbTTG1, LbCPC, and LbTRY (negative regulators) [6]. These genes are components of the MYB–bHLH–WD40 regulatory complex that controls trichome development in model plants such as A. thaliana [21]. The similarity between trichome formation and salt gland development suggests that conserved regulatory modules may underlie epidermal cell differentiation across plant species.
However, it is important to distinguish between transcriptional effects and direct molecular interactions. Current evidence indicates that LbVQ6 influences the expression of these regulatory genes, but there is no direct experimental evidence demonstrating physical interaction between LbVQ6 and LbEGL3, LbTTG1, LbCPC, or LbTRY proteins [22]. Therefore, LbVQ6 is more likely to act as an upstream or intermediate regulator within a broader signaling network rather than as a direct component of the MYB–bHLH–WD40 complex.
The comparison between salt gland development and trichome formation provides a useful conceptual framework; however, this analogy should be interpreted cautiously. While similar regulatory components are involved, the extent to which these pathways are conserved or functionally equivalent in halophytes remains unclear. Additional studies are required to determine whether the regulatory networks controlling salt gland development are directly homologous to those governing trichome differentiation or represent modified or partially conserved pathways.
Another important consideration is that the observed phenotypic and transcriptional changes following LbVQ6 silencing do not necessarily indicate a single linear regulatory pathway. LbVQ6 may function within a complex network integrating multiple signaling pathways, including developmental, hormonal, and stress-responsive signals. Thus, the effects of LbVQ6 on salt gland development may be mediated indirectly through broader transcriptional or signaling cascades rather than through direct gene-specific regulation.
Furthermore, the extent of functional redundancy among VQ family members of L. bicolor remains unknown. Given that multiple VQ genes are present in the genome, it is possible that other VQ proteins contribute to salt gland development either cooperatively or redundantly [23]. The specific roles and interactions of these additional VQ genes have not yet been characterized, representing an important gap in current knowledge.
Taken together, available evidence supports a role for LbVQ6 in regulating salt gland development and associated stress responses; however, its precise molecular function remains incompletely understood. While experimental data clearly demonstrate its involvement in developmental and physiological processes, the underlying regulatory mechanisms-including potential protein interactions, signaling pathways, and downstream targets-require further investigation.
Integration of Hormone and Stress Signaling Pathways
Plant responses to abiotic stress are regulated by complex signaling networks that integrate environmental cues with endogenous hormonal pathways (Figure 2). Phytohormones such as abscisic acid (ABA), jasmonic acid (JA), salicylic acid (SA), and cytokinins play central roles in mediating plant responses to salt stress by regulating gene expression, cellular metabolism, and developmental processes [2,3,24,56]. These hormonal signaling pathways are highly interconnected and often function synergistically or antagonistically to fine-tune stress responses [25,57].
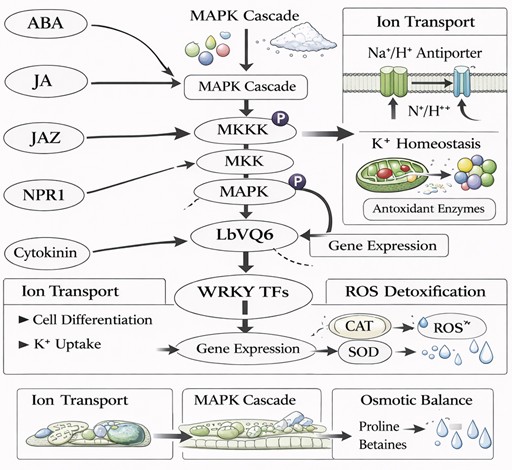
Figure 2. Integration of ABA, JA, SA, and cytokinin signaling pathways mediated by LbVQ6 in regulating salt gland development and salt stress tolerance.
Promoter analysis of LbVQ genes revealed the presence of multiple cis-regulatory elements associated with hormone and stress signaling, including ABA-responsive elements (ABRE), MYB-binding sites, methyl jasmonate (MeJA)-responsive elements, and salicylic acid-responsive elements [6]. While the presence of these motifs suggests potential responsiveness to hormone signaling pathways, it is important to emphasize that such predictions are based on bioinformatic analyses and do not provide direct evidence of functional regulation.
Experimental validation, including hormone treatment assays, reporter gene analysis, and chromatin immunoprecipitation (ChIP)-based approaches, will be required to determine whether these cis-elements are actively involved in regulating LbVQ gene expression in L. bicolor [26]. Therefore, current evidence supports the possibility of hormone-responsive regulation but does not yet establish direct functional involvement of LbVQ6 in specific hormonal pathways.
Despite these limitations, LbVQ6 may be positioned within hormone-mediated signaling networks that coordinate stress responses and developmental processes. ABA is a key regulator of plant responses to drought and salinity, controlling stomatal closure, osmotic balance, and stress-responsive gene expression [2,24]. JA and SA are primarily associated with defense signaling but also contribute to abiotic stress responses through modulation of antioxidant systems and transcriptional regulation [3,25]. Cytokinins regulate cell division and differentiation and may therefore influence developmental processes such as salt gland formation [27].
However, it remains unclear whether LbVQ6 directly participates in these hormones signaling pathways or whether its effects are mediated indirectly through downstream transcriptional networks. At present, there is no direct experimental evidence demonstrating interaction between LbVQ6 and hormone-responsive transcription factors or signaling components. Thus, the proposed role of LbVQ6 in hormone signaling should be considered a working hypothesis rather than a confirmed mechanism.
The integration of hormone signaling pathways with stress-responsive regulatory networks likely enables plants to coordinate developmental processes with environmental adaptation. In the context of salt gland development, hormonal signaling may influence cell differentiation, pattern formation, and functional activity. LbVQ6, as a transcriptional cofactor, may contribute to this integration by modulating transcriptional responses to upstream signals rather than acting as a primary signaling component.
Furthermore, hormone signaling pathways are closely linked with downstream physiological processes such as ion transport regulation and redox homeostasis. ABA signaling regulates ion transporter activity and maintains Na?/K? balance under salt stress, while JA and SA signaling contribute to activation of antioxidant defense systems [2,24,25]. This functional linkage suggests that hormone-mediated signaling may converge on pathways controlling ion homeostasis and oxidative stress responses.
Importantly, these downstream processes-ion transport regulation and redox homeostasis—represent key physiological mechanisms of salt tolerance and are discussed further in Section 5. Thus, any potential role of LbVQ6 in hormone signaling is likely to be functionally connected to these physiological processes rather than acting in isolation.
In addition to hormonal regulation, cross-talk between signaling pathways further increases the complexity of plant stress responses. MAPK signaling cascades, calcium signaling, and transcription factor networks interact with hormone signaling pathways to form integrated regulatory systems [3,28]. LbVQ6 may function as part of this broader network by linking transcriptional regulation with upstream signaling pathways; however, this integrative role remains to be experimentally validated.
Overall, current evidence suggests that LbVQ6 may act as a regulatory node at the interface of hormone and stress signaling pathways. However, its precise molecular function, including potential interactions with hormone-responsive transcription factors and signaling components, remains unresolved and requires further experimental investigation.
Regulation of Ion Homeostasis and Oxidative Stress Signaling
Salt stress imposes severe physiological constraints on plant cells by disrupting ion balance and inducing oxidative stress. Excessive accumulation of sodium ions (Na?) leads to ionic toxicity, membrane destabilization, and inhibition of enzymatic processes, while reduced potassium ion (K?) levels impair cellular metabolism and osmotic regulation [1,2]. In addition to ionic imbalance, salt stress triggers the overproduction of reactive oxygen species (ROS), including superoxide radicals, hydrogen peroxide, and hydroxyl radicals, which cause oxidative damage to proteins, lipids, and nucleic acids [12,29,53].
To survive under such conditions, plants have evolved complex regulatory systems that maintain ion homeostasis and redox balance (Figure 3). Ion transport systems, including Na?/H? antiporters, potassium transporters, and membrane channels, play essential roles in regulating intracellular ion concentrations [2,30]. At the same time, antioxidant defense systems, including enzymatic components such as superoxide dismutase (SOD), catalase (CAT), and peroxidases (POD), as well as non-enzymatic antioxidants, function to detoxify ROS and protect cellular structures from oxidative damage [12,31].
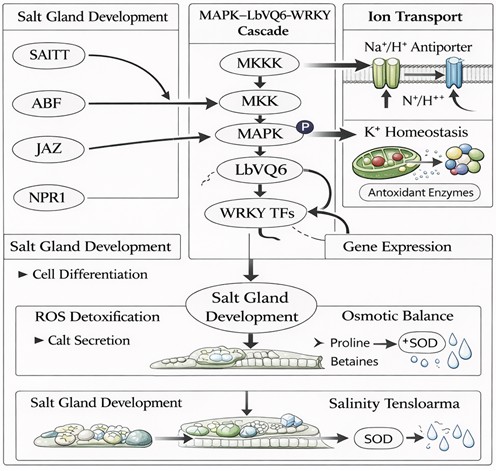
Figure 3. Role of LbVQ6 in regulating ion transport, redox homeostasis, and antioxidant defense signaling pathways under salt stress.
Functional characterization of LbVQ6 has provided important insights into its role in regulating these physiological processes. Gene silencing experiments using virus-induced gene silencing (VIGS), as reported in previous studies, demonstrated that suppression of LbVQ6 expression resulted in significant alterations in ion balance and oxidative stress markers [6,62]. Specifically, LbVQ6-silenced plants exhibited increased Na? accumulation, decreased K? concentration, elevated ROS levels, and enhanced oxidative damage. These observations indicate that LbVQ6 is functionally associated with the maintenance of ion homeostasis and redox balance under salt stress conditions.
However, it is important to distinguish between phenotypic association and direct molecular regulation. The observed changes in ion balance and oxidative stress markers following LbVQ6 silencing do not necessarily demonstrate that LbVQ6 directly regulates ion transporters or antioxidant enzymes. Instead, these effects may be mediated indirectly through broader transcriptional or signaling networks in which LbVQ6 participates.
Based on available evidence, LbVQ6 may influence ion homeostasis by modulating transcriptional pathways that regulate genes involved in ion transport, thereby contributing to the maintenance of Na?/K? balance under saline conditions [30]. Similarly, LbVQ6 may affect antioxidant defense systems through indirect regulation of gene expression controlling ROS detoxification processes [31]. However, these proposed mechanisms remain hypothetical and require experimental validation through transcriptomic, proteomic, and functional analyses.
Importantly, the regulation of ion homeostasis and redox balance is closely linked with upstream signaling pathways, including hormone signaling and MAPK cascades. As discussed in Section 4, hormone signaling pathways such as ABA, JA, and SA are known to regulate ion transport and antioxidant responses [24,25]. LbVQ6 may function as a regulatory intermediary linking these upstream signaling pathways with downstream physiological processes, thereby enabling coordinated stress responses.
The integration of signaling and physiological regulation is essential for plant survival under stress conditions. Efficient ion transport systems prevent toxic ion accumulation, while antioxidant defenses mitigate oxidative damage, together ensuring cellular integrity and metabolic stability. Disruption of these processes, as observed in LbVQ6-silenced plants, highlights the importance of coordinated regulatory networks in stress adaptation rather than the action of a single regulatory component.
In addition, ROS function not only as damaging agents but also as signaling molecules that regulate gene expression and stress-responsive pathways [29,32,52,53]. Controlled ROS production is therefore essential for signaling, whereas excessive accumulation leads to cellular damage. Although LbVQ6 may be involved in maintaining ROS balance, there is currently no direct evidence demonstrating that it specifically regulates ROS signaling pathways. Thus, any proposed role of LbVQ6 in ROS signaling should be considered hypothetical and requires further experimental investigation.
Taken together, current evidence supports a role for LbVQ6 in the regulation of ion homeostasis and oxidative stress responses at the physiological level. However, its precise molecular function, including potential direct targets and regulatory mechanisms, remains unclear and requires further investigation within the context of integrated stress signaling networks.
Interaction with MAPK and WRKY Signaling Cascades
VQ motif-containing proteins are widely recognized as transcriptional regulatory cofactors that function through interaction with transcription factors and signaling kinases. In model plant systems, VQ proteins have been shown to interact with WRKY transcription factors and to be phosphorylated by mitogen-activated protein kinases (MAPKs), thereby linking upstream signaling cascades with downstream transcriptional regulation [3,33,47,48].
MAPK signaling pathways represent a major signal transduction system in plants, mediating responses to environmental stresses such as salinity, drought, and pathogen attack. These cascades involve sequential activation of MAP kinase kinase kinases (MAPKKKs), MAP kinase kinases (MAPKKs), and MAP kinases (MAPKs), ultimately leading to phosphorylation of target proteins and activation of stress-responsive gene expression [3,34]. In A. thaliana, MAPKs such as MPK3 and MPK6 have been shown to phosphorylate VQ proteins, thereby influencing their activity and interaction with WRKY transcription factors [5,33].
Based on these findings, it has been proposed that LbVQ6 may participate in MAPK-mediated signaling pathways in L. bicolor (Figure 4). In this conceptual model, environmental stress signals activate MAPK cascades, which may lead to phosphorylation of VQ proteins, including LbVQ6. Phosphorylated VQ proteins could then influence transcriptional regulation through interaction with WRKY transcription factors, potentially affecting genes involved in ion transport, antioxidant defense, and developmental processes [49].
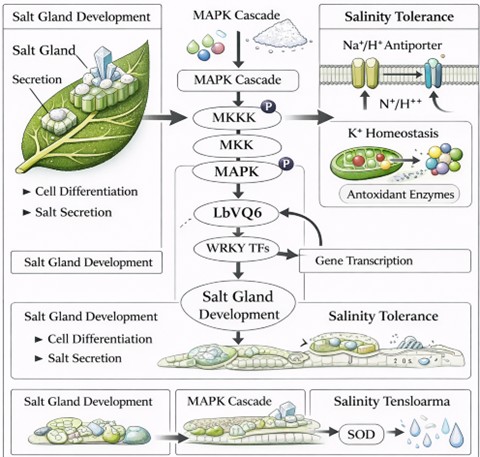
Figure 4. Proposed MAPK–LbVQ6–WRKY signaling cascade regulating salt stress response and salt gland development in L. bicolor.
However, it is critical to emphasize that this MAPK–LbVQ6–WRKY framework remains hypothetical. There is currently no direct experimental evidence demonstrating MAPK-mediated phosphorylation of LbVQ6 or its physical interaction with WRKY transcription factors of L. bicolor. Therefore, the proposed signaling cascade should be interpreted as a model based on functional analogies with other plant systems rather than a confirmed regulatory mechanism.
Furthermore, extrapolation from model species such as A. thaliana to halophytes should be approached with caution. While VQ–WRKY and MAPK–VQ interactions are well documented in model plants, the extent to which these interactions are conserved in L. bicolor remains unclear. Species-specific regulatory differences may exist, particularly given the unique physiological adaptations of halophytes.
In addition, the specific WRKY transcription factors that may interact with LbVQ6 have not yet been identified, and their roles in salt gland development and salt stress responses remain largely unexplored. Similarly, the identity of upstream MAP kinases potentially involved in LbVQ6 phosphorylation has not been determined. These gaps highlight the need for targeted experimental studies, including protein-protein interaction assays, kinase activity analyses, and functional characterization of candidate signaling components [35,47].
Despite these limitations, the MAPK–VQ–WRKY framework provides a useful conceptual basis for understanding how upstream stress signals could be translated into transcriptional responses. Evidence from other plant systems indicates that VQ proteins can function as intermediates linking kinase signaling cascades with transcription factor networks [5,33]. However, whether LbVQ6 performs a similar role in L. bicolor remains to be experimentally validated.
The integration of MAPK signaling with transcription factor networks enables plants to rapidly perceive and respond to environmental stress. Phosphorylation of regulatory proteins such as VQ proteins can alter their activity, stability, and interaction with transcription factors, thereby enabling dynamic regulation of gene expression [34,36]. In this context, LbVQ6 may function as a regulatory intermediary rather than a primary signaling component, although this role has not yet been confirmed.
Furthermore, cross-talk between MAPK signaling and other regulatory pathways, including hormone signaling and redox signaling, contributes to the complexity of plant stress responses. As discussed in previous sections, LbVQ6 may be associated with multiple signaling pathways; however, its precise position within these networks remains unclear [24,32].
Taken together, available evidence suggests that LbVQ6 may be involved in MAPK-related signaling and transcriptional regulation, but the MAPK–LbVQ6–WRKY cascade should be regarded as a hypothesis-driven model. Confirmation of this model will require experimental identification of interaction partners, characterization of phosphorylation events, and functional analysis of downstream target genes [54,55].
Functional Significance in Stress Adaptation and Crop Improvement
Understanding the molecular signaling mechanisms regulated by LbVQ6 provides important insights into plant adaptive responses to salinity and offers potential avenues for improving crop stress tolerance through molecular and genetic approaches. Salinity stress remains a major constraint on global agricultural productivity, affecting a large proportion of cultivated land and significantly reducing crop yield and quality [1,37,63]. Improving plant salt tolerance is therefore a key objective for sustainable agriculture and food security.
Regulatory proteins such as VQ motif-containing transcriptional cofactors play essential roles in coordinating stress-responsive signaling networks and may represent potential targets for crop improvement. Functional studies in L. bicolor have demonstrated that LbVQ6 contributes to salt gland development, ion homeostasis, and oxidative stress tolerance [6,50,55]. These findings suggest that VQ-mediated regulatory pathways could be explored as targets for improving stress tolerance in crop species (Table 2), although their applicability beyond halophytes remains to be established.
|
Application |
Mechanism |
Expected outcome |
Reference |
|
Overexpression of VQ genes |
Enhanced transcriptional regulation of stress-responsive genes |
Improved salt tolerance and stress resistance |
[2] |
|
CRISPR/Cas9-mediated gene editing |
Modification of stress signaling pathways |
Enhanced stress signaling efficiency |
[66] |
|
Molecular breeding |
Selection of stress-tolerant alleles |
Development of stress-resistant crop varieties |
[40] |
|
Manipulation of ion transport regulation |
Improved ion homeostasis |
Increased salinity tolerance |
[30] |
|
Enhancement of antioxidant defense |
Improved ROS detoxification |
Reduced oxidative stress damage |
[53] |
One potential strategy is the heterologous expression of LbVQ6 in glycophytic crops such as rice (O. sativa) or tomato (S. lycopersicum). However, it is important to recognize that halophyte-specific adaptations, such as salt gland-mediated ion secretion, are absent in most crop species. Therefore, the functional effects of LbVQ6 in glycophytes may differ from those observed in L. bicolor, and its role may be limited to modulation of intracellular signaling pathways rather than structural adaptations. Experimental validation in heterologous systems will be required to determine whether LbVQ6 can enhance stress tolerance through conserved regulatory mechanisms [2,38,51].
Genome editing technologies, including CRISPR/Cas systems, provide opportunities for targeted modification of endogenous regulatory networks. Rather than direct transfer of halophyte genes, editing of native VQ genes or associated signaling components in crops may represent a more feasible approach to improving stress tolerance [39,61,64]. However, the success of such strategies depends on a detailed understanding of VQ gene function, regulatory interactions, and network context, which remains incomplete.
Molecular breeding approaches may also benefit from the identification of natural genetic variation in VQ genes associated with stress tolerance. Comparative analysis of VQ gene alleles in stress-tolerant and sensitive genotypes could support marker-assisted selection and genomic breeding strategies [40,61]. Nevertheless, the contribution of individual VQ genes to complex stress tolerance traits is likely to be context-dependent and influenced by interactions with other regulatory pathways.
Importantly, the practical application of these strategies must consider potential trade-offs between stress tolerance and plant growth or yield. Enhanced stress resistance is often associated with reduced growth under non-stress conditions due to resource allocation constraints and regulatory trade-offs [41]. Therefore, manipulation of VQ-mediated signaling pathways should aim to achieve a balance between stress adaptation and growth performance rather than maximizing stress tolerance alone.
It should also be emphasized that current applications remain largely conceptual. Direct experimental evidence demonstrating the effectiveness of LbVQ6-based strategies in crop improvement is currently lacking. Functional validation through gene expression analysis, physiological characterization, and field-based performance evaluation will be necessary to determine their practical utility.
Overall, LbVQ6 and related VQ proteins represent promising but as yet unvalidated targets for improving plant stress tolerance. Their potential application lies primarily in understanding regulatory networks and identifying candidate pathways for manipulation, rather than immediate direct translation into crop improvement systems.
Limitations and Knowledge Gaps
Despite significant progress in understanding the role of LbVQ6 in salt gland development and stress responses, several important limitations and knowledge gaps remain that should be considered when interpreting current findings and designing future research.
First, functional evidence supporting the role of LbVQ6 is primarily based on virus-induced gene silencing (VIGS) experiments. While VIGS is a useful approach for rapid functional analysis, it may produce partial gene suppression and off-target effects, limiting the ability to draw definitive conclusions about gene function [42]. A key unresolved question is whether complete loss-of-function of LbVQ6 would produce phenotypes consistent with those observed in VIGS experiments. Addressing this question will require stable genetic systems, including CRISPR/Cas-mediated knockout lines or transgenic complementation approaches [39,65].
Second, promoter analysis identifying cis-regulatory elements such as ABRE, MeJA-responsive elements, and MYB-binding sites is based on bioinformatic prediction. While these elements suggest potential responsiveness to hormonal signaling pathways, it remains unclear whether LbVQ6 expression is directly regulated by specific hormones under physiological conditions. A critical question is whether these cis-elements are functionally active in vivo and which transcription factors bind to them. Experimental approaches such as hormone treatment assays, reporter gene systems, and chromatin immunoprecipitation (ChIP) analyses will be required to resolve these questions [26,43].
Third, the proposed signaling interactions involving LbVQ6, including its potential phosphorylation by MAP kinases and interaction with WRKY transcription factors, remain hypothetical. A central unresolved issue is whether LbVQ6 is directly phosphorylated by specific MAPKs and whether it physically interacts with defined WRKY proteins in L. bicolor. Addressing these questions will require targeted protein–protein interaction studies, including yeast two-hybrid assays, co-immunoprecipitation, bimolecular fluorescence complementation, and in vitro kinase assays [35,44].
In addition, the transcriptional regulatory role of LbVQ6 remains insufficiently characterized. It is currently unknown which downstream genes are directly regulated by LbVQ6 and whether it acts as a transcriptional activator, repressor, or co-regulator depending on cellular context. Genome-wide identification of LbVQ6 target genes using transcriptomic approaches and DNA-binding or protein interaction analyses represents an important research priority.
Another important gap concerns the functional relationship between LbVQ6 and other members of the VQ gene family in L. bicolor. It remains unclear whether VQ proteins act redundantly, cooperatively, or antagonistically in regulating stress responses. Dissecting the functional specialization and interaction among VQ family members will be essential for understanding the overall regulatory network [45,60].
Furthermore, the evolutionary interpretation of VQ gene family contraction requires additional investigation. Although reduced gene number may be associated with halophytic adaptation, it remains unclear whether this pattern reflects adaptive selection, neutral evolution, or lineage-specific genomic processes [16,18]. Comparative genomic analyses across multiple halophyte species will be necessary to determine whether VQ gene reduction represents a conserved adaptive feature.
Finally, most current knowledge is derived from a limited number of studies focused on a single species. A critical unresolved question is whether the regulatory mechanisms identified for LbVQ6 are conserved across other halophytes or represent species-specific adaptations. Comparative studies involving multiple halophytic and glycophytic species will be required to address this question and to evaluate the broader significance of VQ-mediated signaling networks [46,59].
Addressing these knowledge gaps will be essential for advancing our understanding of LbVQ6-mediated signaling pathways and their role in plant stress adaptation. Future research should prioritize mechanistic validation, identification of direct molecular interactions, and integration of multi-level regulatory data to construct a comprehensive model of VQ-mediated signaling in halophytes.
Conclusion
The functional characterization of LbVQ6 in the recretohalophyte L. bicolor provides valuable insights into the molecular signaling processes associated with plant adaptation to saline environments. Available evidence indicates that LbVQ6 is involved in the regulation of salt gland development, ion homeostasis, and oxidative stress responses, highlighting its contribution to both developmental and physiological aspects of salt tolerance. While current findings suggest that LbVQ6 may participate in coordinating multiple signaling pathways, including hormone-mediated and kinase-associated regulatory networks, its precise molecular role and direct interactions remain to be fully elucidated. Rather than acting as a confirmed central signaling hub, LbVQ6 is more appropriately considered a candidate regulatory component within a complex and integrated stress-response network. These findings improve our understanding of VQ-mediated regulatory systems in halophytes and provide a foundation for future studies aimed at dissecting signaling mechanisms and evaluating their potential relevance for crop stress tolerance.
Acknowledgement
The authors sincerely acknowledge the staff of the Agrobiotechnologies and Biochemistry Scientific Research Institute for their valuable assistance and professional support during the experimental work, biochemical analyses, and interpretation of the results. The authors also gratefully acknowledge Gulistan State University for providing laboratory facilities, technical infrastructure, and a supportive research environment that made this study possible.
CRediT Authorship Contribution Statement
Khabibjon Kushiev: Conceptualization, Writing – original draft, Methodology, Writing-review&editing; Min Chen: Formal analysis, Data curation, Writing – review & editing; Sarvinoz Khojiboboeva: Investigation, Literature analysis, Discussion of the content, Figure design and visualization; Ablakulova Nodira: Investigation, Literature analysis; Abdukarimov Abdulaziz: Formal analysis, Investigation, Literature analysis; Jumonov Utkir, Data curation, Investigation, Literature analysis; Sodikova Sevara: Investigation, Literature analysis; Djuraev Tulkin: Discussion of the content; Tojikulov Abdukhalil: Figure design and visualization; Burkhev Farkhod: Literature analysis, Discussion of the content, Figure design; Khudaynazarov Mukhriddin: Literature analysis, Discussion of the content, Figure design; Sultonova Nigora: Investigation, Methodology
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
2. Zhu JK. Abiotic Stress Signaling and Responses in Plants. Cell. 2016 Oct 6;167(2):313–24.
3. Meng X, Zhang S. MAPK cascades in plant disease resistance signaling. Annu Rev Phytopathol. 2013;51:245–66.
4. Cheng Y, Zhou Y, Yang Y, Chi YJ, Zhou J, Chen JY, et al. Structural and functional analysis of VQ motif-containing proteins in Arabidopsis as interacting proteins of WRKY transcription factors. Plant Physiol. 2012 Jun;159(2):810–25.
5. Pecher P, Eschen-Lippold L, Herklotz S, Kuhle K, Naumann K, Bethke G, et al. The Arabidopsis thaliana mitogen-activated protein kinases MPK3 and MPK6 target a subclass of 'VQ-motif'-containing proteins to regulate immune responses. New Phytol. 2014 Jul;203(2):592–606.
6. Yang R, Zhang Z, Jiang A, Zhu Z, Chen X, Xu H, et al. Genome-wide identification of the VQ gene family from Limonium bicolor and functional characterization of LbVQ6 under salt stress. Plant Physiol Biochem. 2026 Feb;231:110978.
7. Niehl, A., Peña, E.J., Amari, K. and Heinlein, M. (2013), Microtubules in viral replication and transport. Plant J, 75: 290–308.
8. Billakurthi K. A role for the OsMPK6 signaling pathway in regulating rice grain size. Plant Physiol. 2026 Feb 6;200(2):kiaf632
9. Cheng X, Gao C, Liu X, Xu D, Pan X, Gao W, et al. Characterization of the wheat VQ protein family and expression of candidate genes associated with seed dormancy and germination. BMC Plant Biol. 2022 Mar 15;22(1):119.
10. Huang B, Huang Z, Ma R, Ramakrishnan M, Chen J, Zhang Z, et al. Genome-wide identification and expression analysis of LBD transcription factor genes in Moso bamboo (Phyllostachys edulis). BMC Plant Biol. 2021 Jun 28;21(1):296.
11. Tian J, Zhang J, Francis F. The role and pathway of VQ family in plant growth, immunity, and stress response. Planta. 2023 Dec 11;259(1):16.
12. Ahammed GJ, Li Z, Chen J, Dong Y, Qu K, Guo T, et al. Reactive oxygen species signaling in melatonin-mediated plant stress response. Plant Physiol Biochem. 2024 Feb;207:108398.
13. Yuan F, Lyu MJ, Leng BY, Zheng GY, Feng ZT, Li PH, et al. Comparative transcriptome analysis of developmental stages of the Limonium bicolor leaf generates insights into salt gland differentiation. Plant Cell Environ. 2015 Aug;38(8):1637–57.
14. Yuan F, Wang X, Zhao B, Xu X, Shi M, Leng B, et al. The genome of the recretohalophyte Limonium bicolor provides insights into salt gland development and salinity adaptation during terrestrial evolution. Mol Plant. 2022 Jun 6;15(6):1024–44.
15. Albalat R, Cañestro C. Evolution by gene loss. Nat Rev Genet. 2016 Jul;17(7):379–91.
16. Olson MV. When less is more: gene loss as an engine of evolutionary change. The American Journal of Human Genetics. 1999 Jan 1;64(1):18–23
17. Jackson AC, Carine MA, Chapman MA. Genomics of ecological adaptation in Canary Island Descurainia (Brassicaceae) and comparisons with other Brassicaceae. Ecol Evol. 2024 Aug 8;14(8):e70144.
18. Weisman CM, Murray AW, Eddy SR. Many, but not all, lineage-specific genes can be explained by homology detection failure. PLoS biology. 2020 Nov 2;18(11):e3000862.
19. Girardini KN, Olthof AM, Kanadia RN. Introns: the "dark matter" of the eukaryotic genome. Front Genet. 2023 May 16;14:1150212.
20. Han G, Li Y, Yang Z, Wang C, Zhang Y, Wang B. Molecular Mechanisms of Plant Trichome Development. Front Plant Sci. 2022 Jun 1;13:910228.
21. Ramsay NA, Glover BJ. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005 Feb;10(2):63–70.
22. Flowers TJ, Colmer TD. Salinity tolerance in halophytes. New Phytol. 2008;179(4):945–963.
23. Bari R, Jones JD. Role of plant hormones in plant defence responses. Plant Mol Biol. 2009 Mar;69(4):473–88.
24. Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR. Abscisic acid: emergence of a core signaling network. Annu Rev Plant Biol. 2010;61:651–79.
25. Corné M.J. Pieterse, Dieuwertje Van der Does, Christos Zamioudis, Antonio Leon-Reyes, Saskia C.M. Van Wees. 2012. Hormonal Modulation of Plant Immunity. Annu. Rev. Cell Dev. Biol. 28:489–521.
26. O’Malley RC, Huang SS, Song L, Lewsey MG, Bartlett A, Nery JR, et al. Cistrome and epicistrome features shape the regulatory DNA landscape. Cell. 2016 May 19;165(5):1280–92.
27. Sakakibara H. Cytokinins: activity, biosynthesis, and translocation. Annu. Rev. Plant Biol.. 2006 Jun 2;57(1):431–49.
28. Adzigbe J, Frimpong F, Danquah A, Danquah EY, Asante IK, Abebrese SO, et al. The responses and adaptations of rice (Oryza sativa L.) to drought stress: A review. Climate Smart Agriculture. 2025 Aug 19:100080.
29. Apel K, Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol. 2004;55:373–99.
30. Deinlein U, Stephan AB, Horie T, Luo W, Xu G, Schroeder JI. Plant salt-tolerance mechanisms. Trends Plant Sci. 2014 Jun;19(6):371–9.
31. Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem. 2010 Dec;48(12):909–30.
32. Fedoreyeva LI. ROS as Signaling Molecules to Initiate the Process of Plant Acclimatization to Abiotic Stress. Int J Mol Sci. 2024 Nov 4;25(21):11820.
33. Chi Y, Yang Y, Zhou Y, Zhou J, Fan B, Yu JQ, et al. Protein-protein interactions in the regulation of WRKY transcription factors. Mol Plant. 2013 Mar;6(2):287–300.
34. Thilakarathne AS, Liu F, Zou Z. Plant Signaling Hormones and Transcription Factors: Key Regulators of Plant Responses to Growth, Development, and Stress. Plants (Basel). 2025 Mar 31;14(7):1070.
35. Melicher P, Dvořák P, Šamaj J, Takáč T. Protein-protein interactions in plant antioxidant defense. Front Plant Sci. 2022 Dec 14;13:1035573.
36. Rodriguez MC, Petersen M, Mundy J. Mitogen-activated protein kinase signaling in plants. Annu Rev Plant Biol. 2010;61:621–49.
37. FAO. Global status of salt-affected soils—main report. Rome. 2024.
38. Hirayama T, Shinozaki K. Research on plant abiotic stress responses in the post-genome era: past, present and future. Plant J. 2010 Mar;61(6):1041–52.
39. Niraula S, Khanal R, Ghimire N. CRISPR-Cas based precision genome editing: Current advances and associated challenges in crop improvement and trait enhancement. Genetics & Applications. 2024 Dec 24;8(2).
40. Varshney RK, Graner A, Sorrells ME. Genomics-assisted breeding for crop improvement. Trends Plant Sci. 2005 Dec 1;10(12):621–30.
41. Huot B, Yao J, Montgomery BL, He SY. Growth-defense tradeoffs in plants: a balancing act to optimize fitness. Mol Plant. 2014 Aug;7(8):1267–87.
42. He G, Zhao X, Xu Y, Wang Y, Zhang Z, Xiao L, et al. An efficient virus‐induced gene silencing (VIGS) system for gene functional studies in Miscanthus. Gcb Bioenergy. 2023 Jun;15(6):805–20.
43. Kaufmann K, Muiño JM, Østerås M, Farinelli L, Krajewski P, Angenent GC. Chromatin immunoprecipitation (ChIP) of plant transcription factors followed by sequencing (ChIP-SEQ) or hybridization to whole genome arrays (ChIP-CHIP). Nat Protoc. 2010 Mar;5(3):457–72.
44. Strotmann VI, Stahl Y. Visualization of in vivo protein-protein interactions in plants. J Exp Bot. 2022 Jun 24;73(12):3866–80.
45. Panchy N, Lehti-Shiu M, Shiu SH. Evolution of Gene Duplication in Plants. Plant Physiol. 2016 Aug;171(4):2294–316.
46. Rosales-Nieblas AC, Yamada M, Murillo-Amador B, Yamada S. Exploring Salinity Tolerance in Three Halophytic Plants: Physiological and Biochemical Responses to Agronomic Management in a Half-Strength Seawater Aquaponics System. Horticulturae. 2025 Jun 2;11(6):623.
47. Chen F, Hu Y, Vannozzi A, Wu K, Cai H, Qin Y, et al. The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 2017 Nov 2;36(5-6):311–35.
48. Cheng X, Wang Y, Xiong R, Gao Y, Yan H, Xiang Y. A Moso bamboo gene VQ28 confers salt tolerance to transgenic Arabidopsis plants. Planta. 2020 May;251(5):99.
49. Cheng X, Yao H, Cheng Z, Tian B, Gao C, Gao W, et al. The Wheat Gene TaVQ14 Confers Salt and Drought Tolerance in Transgenic Arabidopsis thaliana Plants. Front Plant Sci. 2022 May 10;13:870586.
50. Hu L, Zheng T, Cai M, Pan H, Wang J, Zhang Q. Transcriptome analysis during floral organ development provides insights into stamen petaloidy in Lagerstroemia speciosa. Plant Physiol Biochem. 2019 Sep;142:510–518.
51. Jeffares DC, Penkett CJ, Bähler J. Rapidly regulated genes are intron poor. Trends Genet. 2008 Aug;24(8):375–8.
52. Liu Y, He C. A review of redox signaling and the control of MAP kinase pathway in plants. Redox Biol. 2017 Apr;11:192–204.
53. Hasanuzzaman M, Bhuyan MHMB, Zulfiqar F, Raza A, Mohsin SM, Mahmud JA, et al. Reactive Oxygen Species and Antioxidant Defense in Plants under Abiotic Stress: Revisiting the Crucial Role of a Universal Defense Regulator. Antioxidants (Basel). 2020 Jul 29;9(8):681.
54. Chen G, Deng W, Peng F, Truksa M, Singer S, Snyder CL, et al. Brassica napus TT16 homologs with different genomic origins and expression levels encode proteins that regulate a broad range of endothelium-associated genes at the transcriptional level. Plant J. 2013 May;74(4):663–77.
55. Kim DY, Kwon SI, Choi C, Lee H, Ahn I, Park SR, et al. Expression analysis of rice VQ genes in response to biotic and abiotic stresses. Gene. 2013 Oct 25;529(2):208–14.
56. Kieber JJ, Schaller GE. Cytokinin signaling in plant development. Development. 2018 Feb 27;145(4):dev149344.
57. Klessig DF, Durner J, Noad R, Navarre DA, Wendehenne D, Kumar D, et al. Nitric oxide and salicylic acid signaling in plant defense. Proc Natl Acad Sci U S A. 2000 Aug 1;97(16):8849–55.
58. Murray CS, Bergland AO. Patterns of Gene Family Evolution and Selection Across Daphnia. Ecol Evol. 2025 May 24;15(5):e71453.
59. Velasco R, Zharkikh A, Affourtit J, Dhingra A, Cestaro A, Kalyanaraman A, et al. The genome of the domesticated apple (Malus× domestica Borkh.). Nature genetics. 2010 Oct;42(10):833–9.
60. Chen M, Wang H, Li X, Li N, Xu G, Meng Q. PLIN1 deficiency affects testicular gene expression at the meiotic stage in the first wave of spermatogenesis. Gene. 2014 Jun 15;543(2):212–9.
61. Wang Z, Gerstein M, Snyder M. RNA-Seq: a revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009 Jan;10(1):57–63.
62. Wang Y, Duan X, Xu S, Wang R, Ouyang Z, Shen W. Linking hydrogen-mediated boron toxicity tolerance with improvement of root elongation, water status and reactive oxygen species balance: a case study for rice. Ann. Bot. 2016 Dec 1;118(7):1279–91.
63. Song M, Zhao J, Zhang C, Jia C, Yang J, Zhao H, et al. PEA-m6A: an ensemble learning framework for accurately predicting N 6-methyladenosine modifications in plants. Plant Physiol. 2024 Jun;195(2):1200–13.
64. Zhu JK. Salt and drought stress signal transduction in plants. Annu Rev Plant Biol. 2002;53:247–73.
65. Zhu JK. Regulation of ion homeostasis under salt stress. Curr Opin Plant Biol. 2003 Oct;6(5):441–5.